
Липиды представляют собой широкую группу органических соединений, включающую жиры , воски , стерины , жирорастворимые витамины (такие как витамины A , D , E и K ), моноглицериды , диглицериды , фосфолипиды и другие. Функции липидов включают хранение энергии, передачу сигналов и выполнение функций структурных компонентов клеточных мембран . [3] [4] Липиды находят применение в косметической и пищевой промышленности , а также в нанотехнологиях . [5]
Липиды можно в широком смысле определить как гидрофобные или амфифильные малые молекулы; амфифильная природа некоторых липидов позволяет им образовывать структуры, такие как везикулы , многослойные/ однослойные липосомы или мембраны в водной среде. Биологические липиды полностью или частично происходят из двух различных типов биохимических субъединиц или «строительных блоков»: кетоацильных и изопреновых групп. [3] Используя этот подход, липиды можно разделить на восемь категорий: жирные ацилы , глицеролипиды , глицерофосфолипиды , сфинголипиды , сахаролипиды и поликетиды (полученные в результате конденсации кетоацильных субъединиц); и стероловые липиды и преноловые липиды (полученные в результате конденсации изопреновых субъединиц). [3]
Хотя термин «липид» иногда используется как синоним жиров, жиры являются подгруппой липидов, называемых триглицеридами . Липиды также охватывают такие молекулы, как жирные кислоты и их производные (включая три-, ди-, моноглицериды и фосфолипиды), а также другие стеролсодержащие метаболиты , такие как холестерин . [6] Хотя люди и другие млекопитающие используют различные биосинтетические пути как для расщепления, так и для синтеза липидов, некоторые незаменимые липиды не могут быть получены таким образом и должны быть получены из пищи.
В 1815 году Анри Браконно классифицировал липиды ( graisses ) на две категории: suifs (твердые жиры или сало) и huiles (жидкие масла). [7] В 1823 году Мишель Эжен Шеврель разработал более подробную классификацию, включив масла, смазки, сало, воски, смолы, бальзамы и летучие масла (или эфирные масла). [8] [9] [10]
Первый синтетический триглицерид был описан Теофилем-Жюлем Пелузом в 1844 году, когда он получил трибутирин путем обработки масляной кислоты глицерином в присутствии концентрированной серной кислоты . [11] Несколько лет спустя Марселен Бертело , один из учеников Пелуза, синтезировал тристеарин и трипальмитин путем реакции аналогичных жирных кислот с глицерином в присутствии газообразного хлористого водорода при высокой температуре. [12]
В 1827 году Уильям Праут признал жир («маслянистые» пищевые вещества), наряду с белками («белковыми») и углеводами («сахаристыми»), важными питательными веществами для людей и животных. [13] [14]
В течение столетия химики считали «жиры» только простыми липидами, состоящими из жирных кислот и глицерина (глицериды), но позже были описаны новые формы. Теодор Гобли (1847) открыл фосфолипиды в мозге млекопитающих и куриных яйцах, названные им « лецитинами ». Тудихум открыл в человеческом мозге некоторые фосфолипиды ( цефалин ), гликолипиды ( цереброзид ) и сфинголипиды ( сфингомиелин ). [9]
Термины липоид, липин, липид и липид использовались в разных значениях от автора к автору. [15] В 1912 году Розенблум и Гис предложили заменить «липоид» на «липин». [16] В 1920 году Блур ввел новую классификацию для «липоидов»: простые липоиды (смазки и воски), сложные липоиды (фосфолипиды и гликолипиды) и производные липоиды (жирные кислоты, спирты , стерины). [17] [18]
Слово липид , которое этимологически происходит от греческого λίπος, lipos «жир», было введено в 1923 году французским фармакологом Габриэлем Бертраном . [19] Бертран включил в концепцию не только традиционные жиры (глицериды), но и «липоиды», имеющие сложную конституцию. [9] Слово липид было единогласно одобрено международной комиссией Société de Chimie Biologique на пленарном заседании 3 июля 1923 года. Слово липид позже было англицировано как липид из-за его произношения ('lɪpɪd). Во французском языке суффикс -ide , от древнегреческого -ίδης (что означает «сын» или «потомок»), всегда произносится (ɪd).
В 1947 году Т. П. Хилдич определил «простые липиды» как жиры и воски (истинные воски, стерины, спирты). [20] [ нужна страница ]
Консорциум Lipid MAPS [3] классифицировал липиды на восемь категорий следующим образом:


Жирные ацилы, общий термин для описания жирных кислот, их конъюгатов и производных, представляют собой разнообразную группу молекул, синтезируемых путем удлинения цепи праймера ацетил-КоА с группами малонил-КоА или метилмалонил-КоА в процессе, называемом синтезом жирных кислот . [21] [22] Они состоят из углеводородной цепи , которая заканчивается группой карбоксильной кислоты ; такое расположение придает молекуле полярный , гидрофильный конец и неполярный, гидрофобный конец, который нерастворим в воде. Структура жирной кислоты является одной из самых фундаментальных категорий биологических липидов и обычно используется в качестве строительного блока более структурно сложных липидов. Углеродная цепь, обычно длиной от четырех до 24 атомов углерода, [23] может быть насыщенной или ненасыщенной и может быть присоединена к функциональным группам, содержащим кислород , галогены , азот и серу . Если жирная кислота содержит двойную связь, существует вероятность геометрической изомерии цис или транс , что существенно влияет на конфигурацию молекулы . Цис -двойные связи заставляют цепь жирной кислоты изгибаться, эффект, который усугубляется большим количеством двойных связей в цепи. Три двойные связи в 18-углеродной линоленовой кислоте , наиболее распространенных жирно-ацильных цепях тилакоидных мембран растений , делают эти мембраны очень текучими, несмотря на низкие температуры окружающей среды, [24] а также заставляют линоленовую кислоту давать доминирующие острые пики в спектрах ЯМР 13-C высокого разрешения хлоропластов. Это, в свою очередь, играет важную роль в структуре и функционировании клеточных мембран. [25] : 193–5 Большинство встречающихся в природе жирных кислот имеют цис -конфигурацию, хотя транс- форма существует в некоторых природных и частично гидрогенизированных жирах и маслах. [26]
Примерами биологически важных жирных кислот являются эйкозаноиды , полученные в основном из арахидоновой кислоты и эйкозапентаеновой кислоты , которые включают простагландины , лейкотриены и тромбоксаны . Докозагексаеновая кислота также важна в биологических системах, особенно в отношении зрения. [27] [28] Другие основные классы липидов в категории жирных кислот — это жирные эфиры и жирные амиды. Жирные эфиры включают важные биохимические промежуточные продукты, такие как восковые эфиры , производные кофермента А тиоэфира жирных кислот , производные тиоэфира жирных кислот АСР и карнитины жирных кислот. Жирные амиды включают N-ацилэтаноламины , такие как каннабиноидный нейротрансмиттер анандамид . [29]
Глицеролипиды состоят из моно-, ди- и тризамещенных глицеринов , [30] наиболее известными из которых являются триэфиры жирных кислот глицерина, называемые триглицеридами . Слово «триацилглицерол» иногда используется как синоним «триглицерида». В этих соединениях три гидроксильные группы глицерина этерифицированы, как правило, различными жирными кислотами. Поскольку они функционируют как хранилище энергии, эти липиды составляют большую часть запаса жира в тканях животных. Гидролиз эфирных связей триглицеридов и высвобождение глицерина и жирных кислот из жировой ткани являются начальными этапами метаболизма жира. [31] : 630–1
Дополнительные подклассы глицеролипидов представлены гликозилглицеролами, которые характеризуются наличием одного или нескольких остатков сахара, присоединенных к глицерину через гликозидную связь . Примерами структур этой категории являются дигалактозилдиацилглицеролы, обнаруженные в растительных мембранах [32] и семинолипиды из сперматозоидов млекопитающих . [33]

Глицерофосфолипиды, обычно называемые фосфолипидами (хотя сфингомиелины также классифицируются как фосфолипиды), повсеместно распространены в природе и являются ключевыми компонентами липидного бислоя клеток, [34] а также участвуют в метаболизме и клеточной сигнализации . [35] Нервная ткань (включая мозг) содержит относительно большое количество глицерофосфолипидов, и изменения в их составе были связаны с различными неврологическими расстройствами. [36] Глицерофосфолипиды можно подразделить на отдельные классы, основываясь на природе полярной головной группы в положении sn -3 глицеринового остова у эукариот и эубактерий или в положении sn -1 в случае архебактерий . [37]
Примерами глицерофосфолипидов, обнаруженных в биологических мембранах, являются фосфатидилхолин (также известный как PC, GPCho или лецитин ), фосфатидилэтаноламин (PE или GPEtn) и фосфатидилсерин (PS или GPSer). Помимо того, что они служат основным компонентом клеточных мембран и сайтами связывания для внутри- и межклеточных белков, некоторые глицерофосфолипиды в эукариотических клетках, такие как фосфатидилинозитолы и фосфатидные кислоты, являются либо предшественниками, либо сами по себе являются мембранными вторичными мессенджерами . [31] : 844 Обычно одна или обе эти гидроксильные группы ацилируются длинноцепочечными жирными кислотами, но существуют также алкилсвязанные и 1Z-алкенилсвязанные ( плазмалоген ) глицерофосфолипиды, а также варианты диалкилэфиров в архебактериях. [38]

Сфинголипиды представляют собой сложное семейство соединений [39], которые имеют общую структурную особенность, сфингоидную основу, которая синтезируется de novo из аминокислоты серина и длинноцепочечного жирного ацил-КоА, затем преобразуется в церамиды , фосфосфинголипиды, гликосфинголипиды и другие соединения. Основная сфингоидная основа млекопитающих обычно называется сфингозином . Церамиды (N-ацил-сфингоидные основания) представляют собой основной подкласс производных сфингоидных оснований с амидной жирной кислотой. Жирные кислоты обычно насыщенные или мононенасыщенные с длиной цепи от 16 до 26 атомов углерода. [25] : 421–2
Основными фосфофосфинголипидами млекопитающих являются сфингомиелины (церамидные фосфохолины), [40] тогда как насекомые содержат в основном церамидные фосфоэтаноламины [41] , а грибы имеют фитоцерамидные фосфоинозитолы и головные группы, содержащие маннозу . [42] Гликосфинголипиды представляют собой разнообразное семейство молекул, состоящих из одного или нескольких остатков сахара, связанных через гликозидную связь со сфингоидной основой. Примерами этого являются простые и сложные гликосфинголипиды, такие как цереброзиды и ганглиозиды .
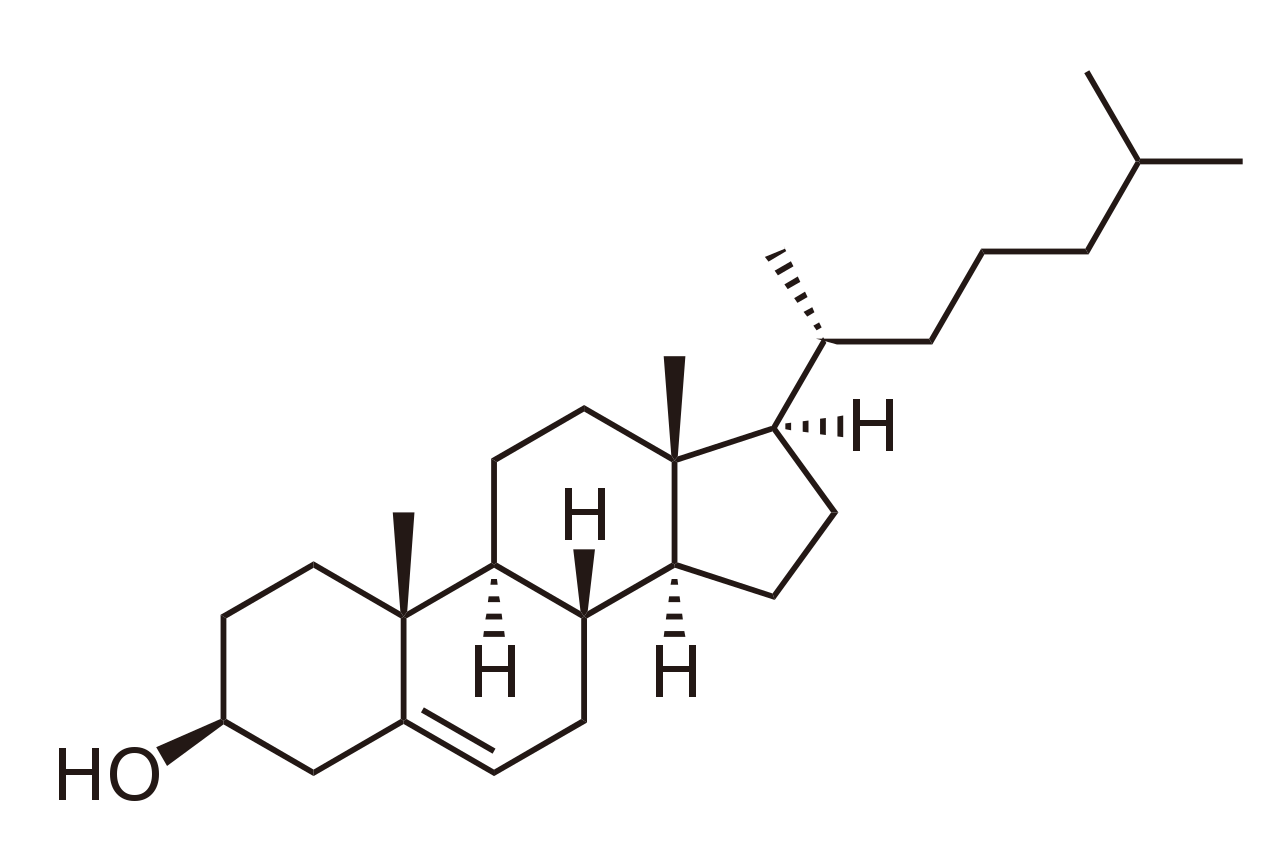
Стерины, такие как холестерин и его производные, являются важным компонентом мембранных липидов, [43] наряду с глицерофосфолипидами и сфингомиелинами. Другими примерами стеринов являются желчные кислоты и их конъюгаты, [44] которые у млекопитающих являются окисленными производными холестерина и синтезируются в печени. Растительными эквивалентами являются фитостерины , такие как β-ситостерин , стигмастерин и брассикастерин ; последнее соединение также используется в качестве биомаркера роста водорослей . [45] Преобладающим стеролом в мембранах клеток грибов является эргостерин . [46]
Стерины — это стероиды , в которых один из атомов водорода замещен гидроксильной группой в положении 3 в углеродной цепи. Они имеют общую со стероидами структуру ядра из четырех слитых колец. Стероиды играют различную биологическую роль в качестве гормонов и сигнальных молекул . Стероиды с восемнадцатью атомами углерода (C18) включают семейство эстрогенов , тогда как стероиды с C19 включают андрогены , такие как тестостерон и андростерон . Подкласс C21 включает прогестагены , а также глюкокортикоиды и минералокортикоиды . [2] : 749 Секостероиды , включающие различные формы витамина D , характеризуются расщеплением кольца B структуры ядра. [47]

Преноловые липиды синтезируются из пятиуглеродных предшественников изопентенилдифосфата и диметилаллилдифосфата , которые производятся в основном через путь мевалоновой кислоты (МВА). [48] Простые изопреноиды (линейные спирты, дифосфаты и т. д.) образуются путем последовательного добавления единиц С5 и классифицируются в соответствии с числом этих терпеновых единиц. Структуры, содержащие более 40 атомов углерода, известны как политерпены. Каротиноиды являются важными простыми изопреноидами, которые функционируют как антиоксиданты и как предшественники витамина А. [ 49] Другой биологически важный класс молекул представлен хинонами и гидрохинонами , которые содержат изопреноидный хвост, прикрепленный к хиноидному ядру неизопреноидного происхождения. [50] Витамин Е и витамин К , а также убихиноны являются примерами этого класса. Прокариоты синтезируют полипренолы (называемые бактопренолами ), в которых конечная изопреноидная единица, присоединенная к кислороду, остается ненасыщенной, тогда как в животных полипренолах ( долихолах ) конечная изопреноидная единица восстановлена. [51]

Сахаролипиды описывают соединения, в которых жирные кислоты связаны с сахарным остовом, образуя структуры, совместимые с мембранными бислоями. В сахаролипидах моносахарид заменяет глицериновый остов, присутствующий в глицеролипидах и глицерофосфолипидах. Наиболее известные сахаролипиды — это ацилированные глюкозаминовые предшественники липида А, компонента липополисахаридов в грамотрицательных бактериях . Типичные молекулы липида А — это дисахариды глюкозамина, которые дериватизированы семью жирно-ацильными цепями. Минимальный липополисахарид, необходимый для роста E. coli , — это Kdo 2 -липид A, гексаацилированный дисахарид глюкозамина, который гликозилирован двумя остатками 3-дезокси-D-манно-октулозоновой кислоты (Kdo). [52]
Поликетиды синтезируются путем полимеризации ацетильных и пропионильных субъединиц классическими ферментами, а также итеративными и многомодульными ферментами, которые разделяют механистические особенности с синтазы жирных кислот . Они включают в себя множество вторичных метаболитов и натуральных продуктов из животных, растительных, бактериальных, грибковых и морских источников и имеют большое структурное разнообразие. [53] [54] Многие поликетиды представляют собой циклические молекулы, остовы которых часто дополнительно модифицируются гликозилированием , метилированием , гидроксилированием , окислением или другими процессами. Многие обычно используемые противомикробные , противопаразитарные и противораковые агенты представляют собой поликетиды или производные поликетидов, такие как эритромицины , тетрациклины , авермектины и противоопухолевые эпотилоны . [55]
Эукариотические клетки характеризуются компартментализированными мембраносвязанными органеллами , которые выполняют различные биологические функции. Глицерофосфолипиды являются основным структурным компонентом биологических мембран , как клеточная плазматическая мембрана и внутриклеточные мембраны органелл; в клетках животных плазматическая мембрана физически отделяет внутриклеточные компоненты от внеклеточной среды. [ необходима цитата ] Глицерофосфолипиды являются амфипатическими молекулами (содержащими как гидрофобные, так и гидрофильные области), которые содержат глицериновое ядро, связанное с двумя жирнокислотными «хвостами» эфирными связями и с одной «головной» группой фосфатной эфирной связью. [ необходима цитата ] Хотя глицерофосфолипиды являются основным компонентом биологических мембран , другие неглицеридные липидные компоненты, такие как сфингомиелин и стерины (в основном холестерин в мембранах животных клеток), также встречаются в биологических мембранах. [56] [2] : 329–331 В растениях и водорослях галактозилдиацилглицерины [57] и сульфохиновозилдиацилглицерины [32] , в которых отсутствует фосфатная группа, являются важными компонентами мембран хлоропластов и связанных с ними органелл и относятся к наиболее распространенным липидам в фотосинтетических тканях, включая ткани высших растений, водорослей и некоторых бактерий. [58]
Растительные тилакоидные мембраны имеют самый большой липидный компонент небислойного моногалактозилдиглицерида (МГДГ) и мало фосфолипидов; несмотря на этот уникальный липидный состав, было показано, что тилакоидные мембраны хлоропластов содержат динамическую липидно-бислойную матрицу, как было обнаружено с помощью магнитного резонанса и электронного микроскопа. [59]

Биологическая мембрана представляет собой форму ламеллярного фазового липидного бислоя . Образование липидных бислоев является энергетически предпочтительным процессом, когда глицерофосфолипиды, описанные выше, находятся в водной среде. [2] : 333–4 Это известно как гидрофобный эффект . В водной системе полярные головки липидов выравниваются по направлению к полярной водной среде, в то время как гидрофобные хвосты минимизируют свой контакт с водой и имеют тенденцию группироваться вместе, образуя везикулу ; в зависимости от концентрации липида это биофизическое взаимодействие может приводить к образованию мицелл , липосом или липидных бислоев . Также наблюдаются другие агрегации, которые являются частью полиморфизма поведения амфифилов (липидов). Фазовое поведение является областью изучения в рамках биофизики . [60] [61] Мицеллы и бислои образуются в полярной среде в результате процесса, известного как гидрофобный эффект. [62] При растворении липофильного или амфифильного вещества в полярной среде полярные молекулы (т.е. вода в водном растворе) становятся более упорядоченными вокруг растворенного липофильного вещества, поскольку полярные молекулы не могут образовывать водородные связи с липофильными областями амфифила. Таким образом, в водной среде молекулы воды образуют упорядоченную « клатратную » клетку вокруг растворенной липофильной молекулы. [63]
Формирование липидов в мембранах протоклеток представляет собой ключевой шаг в моделях абиогенеза , происхождения жизни. [64]
Триглицериды, хранящиеся в жировой ткани, являются основной формой хранения энергии как у животных, так и у растений. Они являются основным источником энергии при аэробном дыхании. Полное окисление жирных кислот высвобождает около 38 кДж/г (9 ккал/г ), по сравнению с всего лишь 17 кДж/г (4 ккал/г) для окислительного распада углеводов и белков . Адипоцит , или жировая клетка, предназначена для непрерывного синтеза и распада триглицеридов у животных, причем распад контролируется в основном активацией гормонально-чувствительного фермента липазы . [65] Перелетные птицы, которые должны летать на большие расстояния без еды, используют триглицериды в качестве топлива для своих полетов. [2] : 619
Появились доказательства, показывающие, что липидная сигнализация является жизненно важной частью клеточной сигнализации . [66] [67] [68] [69] Липидная сигнализация может происходить посредством активации рецепторов, связанных с G-белком , или ядерных рецепторов , и представители нескольких различных категорий липидов были идентифицированы как сигнальные молекулы и клеточные мессенджеры . [70] К ним относятся сфингозин-1-фосфат , сфинголипид, полученный из церамида, который является мощной молекулой-мессенджером, участвующей в регуляции мобилизации кальция, [71] роста клеток и апоптоза; [72] диацилглицерол и фосфатидилинозитолфосфаты (PIP), участвующие в опосредованной кальцием активации протеинкиназы C ; [73] простагландины , которые являются одним из типов эйкозаноидов, полученных из жирных кислот, участвующих в воспалении и иммунитете ; [74] стероидные гормоны, такие как эстроген , тестостерон и кортизол , которые модулируют множество функций, таких как воспроизводство, метаболизм и кровяное давление; и оксистерины, такие как 25-гидроксихолестерин, которые являются агонистами рецептора X печени . [75] Известно, что фосфатидилсериновые липиды участвуют в передаче сигналов для фагоцитоза апоптотических клеток или частей клеток. Они достигают этого, подвергаясь воздействию внеклеточной поверхности клеточной мембраны после инактивации флиппаз , которые помещают их исключительно на цитозольную сторону, и активации скрамблаз, которые перемешивают ориентацию фосфолипидов. После этого другие клетки распознают фосфатидилсерины и фагоцитируют клетки или фрагменты клеток, подвергая их воздействию. [76]
«Жирорастворимые» витамины ( A , D , E и K ), которые являются липидами на основе изопрена , являются важными питательными веществами, хранящимися в печени и жировых тканях, с разнообразным спектром функций. Ацилкарнитины участвуют в транспорте и метаболизме жирных кислот в митохондриях и из них, где они подвергаются бета-окислению . [77] Полипренолы и их фосфорилированные производные также играют важную транспортную роль, в данном случае транспорт олигосахаридов через мембраны. Полипренолфосфатные сахара и полипренолдифосфатные сахара функционируют в реакциях внецитоплазматического гликозилирования, во внеклеточном биосинтезе полисахаридов (например, полимеризация пептидогликана у бактерий) и в N- гликозилировании эукариотических белков . [78] [79] Кардиолипины представляют собой подкласс глицерофосфолипидов, содержащих четыре ацильные цепи и три глицериновые группы, которые особенно распространены во внутренней митохондриальной мембране. [80] [81] Считается, что они активируют ферменты, участвующие в окислительном фосфорилировании . [82] Липиды также составляют основу стероидных гормонов. [83]
Основными пищевыми липидами для людей и других животных являются животные и растительные триглицериды, стерины и мембранные фосфолипиды. Процесс липидного метаболизма синтезирует и разрушает липидные запасы и производит структурные и функциональные липиды, характерные для отдельных тканей.
У животных при переизбытке диетических углеводов избыток углеводов преобразуется в триглицериды. Это включает синтез жирных кислот из ацетил-КоА и этерификацию жирных кислот при производстве триглицеридов, процесс, называемый липогенезом . [2] : 634 Жирные кислоты производятся синтазы жирных кислот , которые полимеризуют, а затем восстанавливают единицы ацетил-КоА. Ацильные цепи в жирных кислотах удлиняются циклом реакций, которые добавляют ацетильную группу, восстанавливают ее до спирта, дегидратируют ее до алкеновой группы, а затем снова восстанавливают ее до алкановой группы. Ферменты биосинтеза жирных кислот делятся на две группы: у животных и грибов все эти реакции синтазы жирных кислот осуществляются одним многофункциональным белком, [84] в то время как в растительных пластидах и бактериях отдельные ферменты выполняют каждый шаг в пути. [85] [86] Жирные кислоты впоследствии могут быть преобразованы в триглицериды, которые упаковываются в липопротеины и секретируются из печени.
Синтез ненасыщенных жирных кислот включает реакцию десатурации , в ходе которой в жирную ацильную цепь вводится двойная связь. Например, у людей десатурация стеариновой кислоты стеароил -КоА-десатуразой-1 приводит к образованию олеиновой кислоты . Дважды ненасыщенная жирная кислота линолевая кислота , а также трижды ненасыщенная α-линоленовая кислота не могут быть синтезированы в тканях млекопитающих, и поэтому являются незаменимыми жирными кислотами и должны поступать с пищей. [2] : 643
Синтез триглицеридов происходит в эндоплазматическом ретикулуме посредством метаболических путей, в которых ацильные группы в жирных ацил-КоА переносятся в гидроксильные группы глицерол-3-фосфата и диацилглицерина. [2] : 733–9
Терпены и изопреноиды , включая каротиноиды , производятся путем сборки и модификации изопреновых единиц, полученных из реакционноспособных предшественников изопентенилпирофосфата и диметилаллилпирофосфата . [48] Эти предшественники могут быть получены разными способами. У животных и архей мевалонатный путь производит эти соединения из ацетил-КоА, [87] тогда как у растений и бактерий немевалонатный путь использует пируват и глицеральдегид-3-фосфат в качестве субстратов. [48] [88] Одной из важных реакций, в которой используются эти активированные доноры изопрена, является биосинтез стероидов . Здесь изопреновые единицы соединяются вместе, образуя сквален , а затем складываются и формируются в набор колец, образуя ланостерол . [89] Затем ланостерол может быть преобразован в другие стероиды, такие как холестерин и эргостерол. [89] [90]
Бета-окисление — это метаболический процесс, при котором жирные кислоты расщепляются в митохондриях или пероксисомах с образованием ацетил-КоА . По большей части жирные кислоты окисляются с помощью механизма, который похож, но не идентичен обратному процессу синтеза жирных кислот. То есть двухуглеродные фрагменты последовательно удаляются с карбоксильного конца кислоты после стадий дегидрирования , гидратации и окисления с образованием бета-кетокислоты , которая расщепляется тиолизом . Затем ацетил-КоА в конечном итоге преобразуется в аденозинтрифосфат (АТФ), CO2 и H2O с использованием цикла лимонной кислоты и цепи переноса электронов . Следовательно, цикл лимонной кислоты может начинаться с ацетил-КоА, когда жир расщепляется для получения энергии, если глюкозы мало или она отсутствует. Выход энергии при полном окислении пальмитата жирной кислоты составляет 106 АТФ. [2] : 625–6 Ненасыщенные и нечетноцепочечные жирные кислоты требуют дополнительных ферментативных стадий для расщепления.
Большая часть жира, содержащегося в пище, находится в форме триглицеридов, холестерина и фосфолипидов. Некоторое количество пищевого жира необходимо для облегчения усвоения жирорастворимых витаминов ( A , D , E и K ) и каротиноидов . [91] : 903 Люди и другие млекопитающие имеют диетическую потребность в определенных незаменимых жирных кислотах, таких как линолевая кислота ( жирная кислота омега-6 ) и альфа-линоленовая кислота (жирная кислота омега-3), поскольку они не могут быть синтезированы из простых предшественников в рационе. [2] : 643 Обе эти жирные кислоты являются 18-углеродными полиненасыщенными жирными кислотами, различающимися по количеству и положению двойных связей. Большинство растительных масел богаты линолевой кислотой ( масла сафлора , подсолнечника и кукурузы ). Альфа-линоленовая кислота содержится в зеленых листьях растений и в некоторых семенах, орехах и бобовых (в частности , льне , рапсе , грецком орехе и сое ). [92] Рыбий жир особенно богат длинноцепочечными омега-3 жирными кислотами: эйкозапентаеновой кислотой и докозагексаеновой кислотой . [91] : 388 Многие исследования показали положительные преимущества для здоровья, связанные с потреблением омега-3 жирных кислот, в отношении развития младенцев, рака, сердечно-сосудистых заболеваний и различных психических заболеваний (таких как депрессия, синдром дефицита внимания и гиперактивности и деменция). [93] [94]
Напротив, в настоящее время общеизвестно, что потребление трансжиров , таких как те, которые присутствуют в частично гидрогенизированных растительных маслах , является фактором риска сердечно-сосудистых заболеваний . Жиры, которые полезны для человека, могут быть превращены в трансжиры неправильными методами приготовления пищи, которые приводят к пережариванию липидов. [95] [96] [97]
Несколько исследований предположили, что общее потребление жиров в рационе связано с повышенным риском ожирения. [98] [99] и диабета; [100] Другие, включая исследование по изменению рациона питания Women's Health Initiative, восьмилетнее исследование 49 000 женщин, исследование Nurses' Health Study и исследование Health Professionals Follow-up Study, не выявили таких связей. [101] [102] Ни одно из этих исследований не предположило никакой связи между процентом калорий из жиров и риском рака, сердечных заболеваний или увеличения веса. The Nutrition Source, [103] веб-сайт, поддерживаемый кафедрой питания в Школе общественного здравоохранения TH Chan в Гарвардском университете , суммирует текущие доказательства влияния жиров в рационе: «Подробные исследования, большая часть которых проводилась в Гарварде, показывают, что общее количество жиров в рационе на самом деле не связано с весом или заболеванием». [104]
Вводный
Номенклатура
Базы данных
Общий
{kind=link}