Губки или морские губки являются представителями многоклеточного типа Porifera [4] ( / p ə ˈ r ɪ f ər ə ˌ p ɔː - / pər- IF -ər-ə, por- ; означает «носитель пор»), [5] базальной группы животных и родственного таксона диплобластов . [6] Они являются сидячими фильтраторами , которые привязаны к морскому дну , и являются одними из древнейших представителей макробентоса , причем многие исторические виды являются важными рифообразующими организмами.
Губки — многоклеточные организмы, состоящие из желеобразного мезогила, зажатого между двумя тонкими слоями клеток , и обычно имеющие трубчатые тела, полные пор и каналов, которые позволяют воде циркулировать через них. У них есть неспециализированные клетки, которые могут трансформироваться в другие типы и которые часто мигрируют между основными слоями клеток и мезогилом в этом процессе. У них нет сложной нервной , [7] пищеварительной или кровеносной систем . Вместо этого большинство из них полагаются на поддержание постоянного потока воды через свои тела для получения пищи и кислорода и удаления отходов, обычно с помощью движений жгутиков так называемых « воротничковых клеток ».
Считается, что губки являются одними из самых базальных животных, живущих сегодня, и, возможно, были первой внешней группой, ответвившейся от эволюционного древа от последнего общего предка всех животных , [6] с ископаемыми свидетельствами примитивных губок, таких как Otavia, начиная с тонийского периода (около 800 млн лет назад ). Раздел зоологии , изучающий губки, известен как спонгиология . [8]
Термин «губка» происходит от древнегреческого слова σπόγγος spóngos . [9] Научное название Porifera является средним родом множественного числа современного латинского термина porifer , который происходит от корней porus , означающих «пора, отверстие», и -fer, означающих «несущий или переносящий».

Губки похожи на других животных тем, что они многоклеточные , гетеротрофные , не имеют клеточных стенок и производят сперматозоиды . В отличие от других животных, у них нет настоящих тканей [10] и органов . [11] Некоторые из них радиально симметричны, но большинство асимметричны. Формы их тел адаптированы для максимальной эффективности потока воды через центральную полость, где вода откладывает питательные вещества, а затем выходит через отверстие, называемое оскулюм . Одноклеточные хоанофлагелляты напоминают клетки хоаноцитов губок, которые используются для управления их системами потока воды и захвата большей части их пищи. Это наряду с филогенетическими исследованиями рибосомных молекул было использовано в качестве морфологического доказательства, чтобы предположить, что губки являются сестринской группой по отношению к остальным животным. [12] Подавляющее большинство из них являются морскими (солеными) видами, среда обитания которых варьируется от приливных зон до глубин, превышающих 8800 м (5,5 миль), хотя есть и пресноводные виды. Все взрослые губки сидячие , то есть они прикрепляются к подводной поверхности и остаются на месте (т. е. не перемещаются). На личиночной стадии они подвижны .
Многие губки имеют внутренние скелеты из спикул (скелетоподобных фрагментов карбоната кальция или диоксида кремния ) и/или спонгина (модифицированного типа коллагенового белка). [10] Внутренний желатиновый матрикс , называемый мезогил, функционирует как эндоскелет , и это единственный скелет в мягких губках, который покрывает такие твердые поверхности, как камни. Чаще всего мезогил укреплен минеральными спикулами , волокнами спонгина или и тем, и другим. 90% всех известных видов губок, имеющих самый широкий спектр местообитаний, включая все пресноводные, являются демоспонгами , которые используют спонгин; у многих видов есть кремниевые спикулы, тогда как у некоторых видов есть экзоскелеты из карбоната кальция . Известковые губки имеют спикулы из карбоната кальция, а у некоторых видов — экзоскелеты из карбоната кальция, ограничены относительно мелководными морскими водами, где проще всего производить карбонат кальция. [13] : 179 Хрупкие стеклянные губки с « лесами » из кремниевых спикул ограничены полярными регионами и глубинами океана, где хищники редки. Окаменелости всех этих типов были найдены в породах, датируемых 580 миллионами лет назад . Кроме того, археоциаты , окаменелости которых обычны в породах возрастом от 530 до 490 миллионов лет назад , теперь рассматриваются как тип губки.
Хотя большинство из приблизительно 5000–10000 известных видов губок питаются бактериями и другой микроскопической пищей в воде, некоторые из них содержат фотосинтезирующие микроорганизмы в качестве эндосимбионтов , и эти союзы часто производят больше пищи и кислорода, чем потребляют. Несколько видов губок, которые живут в среде с недостатком пищи, эволюционировали как плотоядные , которые охотятся в основном на мелких ракообразных . [14]
Большинство губок размножаются половым путем , но они также могут размножаться бесполым путем. Виды, размножающиеся половым путем, выпускают сперматозоиды в воду, чтобы оплодотворить яйцеклетки, выпущенные или сохраненные их партнером или «матерью»; оплодотворенные яйца развиваются в личинок , которые уплывают в поисках мест для поселения. [13] : 183–185 Известно, что губки регенерируют из фрагментов, которые отламываются, хотя это работает только в том случае, если фрагменты включают правильные типы клеток. Некоторые виды размножаются почкованием. Когда условия окружающей среды становятся менее благоприятными для губок, например, при падении температуры, многие пресноводные виды и несколько морских производят геммулы , «стручки выживания» неспециализированных клеток, которые остаются в состоянии покоя, пока условия не улучшатся; затем они либо образуют совершенно новые губки, либо повторно колонизируют скелеты своих родителей. [13] : 120–127

Несколько видов демоспонга, которые имеют полностью мягкие волокнистые скелеты без жестких элементов, использовались людьми на протяжении тысяч лет для различных целей, в том числе в качестве набивки и инструментов для чистки. Однако к 1950-м годам они были настолько истощены , что отрасль почти рухнула, и большинство материалов, похожих на губки, теперь являются синтетическими. Губки и их микроскопические эндосимбионты в настоящее время исследуются как возможные источники лекарств для лечения широкого спектра заболеваний. Было замечено, что дельфины используют губки в качестве инструментов во время добычи пищи . [16]
Губки составляют тип Porifera и определяются как сидячие метазоа (многоклеточные неподвижные животные), у которых отверстия для забора и вывода воды соединены камерами, выстланными хоаноцитами , клетками с кнутообразными жгутиками. [13] : 29 Однако некоторые плотоядные губки утратили эти системы тока воды и хоаноциты. [13] : 39 [17] Все известные живые губки могут перестраивать свои тела, поскольку большинство типов их клеток могут перемещаться внутри их тел, а некоторые могут переходить из одного типа в другой. [17] [18]
Даже если несколько губок способны вырабатывать слизь, которая действует как микробный барьер у всех других животных, не было зарегистрировано ни одной губки со способностью секретировать функциональный слой слизи. Без такого слоя слизи их живая ткань покрыта слоем микробных симбионтов, которые могут составлять до 40–50% влажной массы губки. Эта неспособность предотвратить проникновение микробов в их пористую ткань может быть основной причиной того, что они никогда не развивали более сложную анатомию. [19]
Подобно книдариям (медузы и т. д.) и гребневикам (гребневики), и в отличие от всех других известных многоклеточных животных, тела губок состоят из неживой желеобразной массы ( мезогила ), зажатой между двумя основными слоями клеток. [20] [21] Книдарии и гребневики имеют простую нервную систему, а их клеточные слои связаны внутренними связями и закреплены на базальной мембране (тонкий волокнистый мат, также известный как « базальная пластинка »). [21] У губок нет нервной системы, похожей на нервную систему позвоночных, но может быть совершенно другая. [7] Их средние желеобразные слои имеют большие и разнообразные популяции клеток, и некоторые типы клеток в их внешних слоях могут перемещаться в средний слой и менять свои функции. [18]
Тело губки полое и удерживается в форме мезогилом , желеобразным веществом, состоящим в основном из коллагена и укрепленным плотной сетью волокон, также состоящих из коллагена. Было идентифицировано 18 различных типов клеток. [24] Внутренняя поверхность покрыта хоаноцитами , клетками с цилиндрическими или коническими воротниками, окружающими один жгутик на хоаноцит. Волнообразное движение кнутообразных жгутиков гонит воду через тело губки. Все губки имеют устья , каналы, ведущие внутрь через мезогил, и у большинства губок они контролируются трубчатыми пороцитами , которые образуют закрывающиеся впускные клапаны. Пинакоциты , пластинчатые клетки, образуют однослойную внешнюю оболочку над всеми остальными частями мезогила, которые не покрыты хоаноцитами, и пинакоциты также переваривают частицы пищи, которые слишком велики, чтобы попасть в отверстия, [18] [20] в то время как те, что находятся у основания животного, отвечают за его закрепление. [20]
Другие типы клеток живут и движутся внутри мезогила: [18] [20]
Многие личинки губок обладают глазами без нейронов , которые основаны на криптохромах . Они опосредуют фототаксическое поведение. [25]
Стеклянные губки представляют собой отличительную вариацию этого базового плана. Их спикулы, которые сделаны из кремния , образуют каркас, похожий на леса, между прутьями которого живая ткань подвешена как паутина , содержащая большинство типов клеток. [18] Эта ткань представляет собой синцитий , который в некоторых отношениях ведет себя как множество клеток, которые разделяют одну внешнюю мембрану , а в других — как одна клетка с несколькими ядрами .
Большинство губок работают скорее как дымоходы : они вбирают воду снизу и выбрасывают ее из оскулюма наверху . Поскольку окружающие потоки быстрее наверху, эффект всасывания, который они производят по принципу Бернулли, делает часть работы бесплатно. Губки могут контролировать поток воды с помощью различных комбинаций полного или частичного закрытия оскулюма и устий (впускных пор) и изменения ритма жгутиков, и могут закрыть его, если в воде много песка или ила. [18]
Хотя слои пинакоцитов и хоаноцитов напоминают эпителий более сложных животных, они не связаны прочно межклеточными связями или базальной пластинкой (тонким волокнистым слоем под ними). Гибкость этих слоев и перестройка мезогила лофоцитами позволяют животным изменять свою форму на протяжении всей жизни, чтобы максимально использовать местные водные потоки. [18] : 83
Простейшая структура тела губок — это трубчатая или вазообразная форма, известная как «асконоид», но это сильно ограничивает размер животного. Структура тела характеризуется стеблевидным спонгоцелем, окруженным одним слоем хоаноцитов. Если его просто увеличить, отношение его объема к площади поверхности увеличивается, поскольку поверхность увеличивается как квадрат длины или ширины, а объем увеличивается пропорционально кубу. Количество ткани, которая нуждается в пище и кислороде , определяется объемом, но насосная мощность, которая поставляет пищу и кислород, зависит от площади, покрытой хоаноцитами. Асконоидные губки редко превышают 1 мм (0,039 дюйма) в диаметре. [18]

Некоторые губки преодолевают это ограничение, принимая «сиконоидную» структуру, в которой стенка тела складчатая . Внутренние карманы складок выстланы хоаноцитами, которые соединяются с внешними карманами складок посредством устьиц. Это увеличение числа хоаноцитов и, следовательно, насосной способности позволяет сиконоидным губкам вырастать до нескольких сантиметров в диаметре.
«Лейконоидный» паттерн еще больше увеличивает производительность насоса, заполняя внутреннюю часть почти полностью мезогилом, который содержит сеть камер, выстланных хоаноцитами и соединенных друг с другом и с водозаборами и водовыпусками трубками. Лейконидные губки вырастают до более чем 1 м (3,3 фута) в диаметре, и тот факт, что рост в любом направлении увеличивает количество камер хоаноцитов, позволяет им принимать более широкий спектр форм, например, «корковые» губки, формы которых следуют за формами поверхностей, к которым они прикрепляются. Все пресноводные и большинство мелководных морских губок имеют тела лейконид. Сети водных проходов в стеклянных губках похожи на структуру лейконид. [18]
Во всех трех типах структуры площадь поперечного сечения областей, выстланных хоаноцитами, намного больше, чем у входных и выходных каналов. Это замедляет поток вблизи хоаноцитов и, таким образом, облегчает им захват частиц пищи. [18] Например, в Leuconia , небольшой лейконоидной губке высотой около 10 сантиметров (3,9 дюйма) и диаметром 1 сантиметр (0,39 дюйма), вода поступает в каждый из более чем 80 000 входных каналов со скоростью 6 см в минуту . Однако, поскольку Leuconia имеет более 2 миллионов жгутиконосных камер, общий диаметр которых намного больше диаметра каналов, поток воды через камеры замедляется до 3,6 см в час , что позволяет хоаноцитам легко захватывать пищу. Вся вода выталкивается через один оскулюм со скоростью около 8,5 см в секунду , что достаточно быстро, чтобы унести отходы на некоторое расстояние. [27]

В зоологии скелет — это любая достаточно жесткая структура животного, независимо от того, имеет ли она суставы и является ли она биоминерализованной . Мезохил функционирует как эндоскелет у большинства губок и является единственным скелетом у мягких губок, которые покрывают твердые поверхности, такие как камни. Чаще всего мезогил укреплен минеральными спикулами , волокнами спонгина или обоими. Спикулы, которые присутствуют у большинства, но не у всех видов, [28] могут быть сделаны из кремнезема или карбоната кальция и различаться по форме от простых стержней до трехмерных «звезд» с шестью лучами. Спикулы производятся клетками склероцитов [ 18] и могут быть отдельными, соединенными суставами или слитыми. [17]
Некоторые губки также выделяют экзоскелеты , которые полностью лежат снаружи их органических компонентов. Например, склероспонги («твердые губки») имеют массивные экзоскелеты из карбоната кальция, над которыми органическое вещество образует тонкий слой с камерами хоаноцитов в углублениях в минерале. Эти экзоскелеты выделяются пинакоцитами , которые формируют кожу животных. [18]

Хотя взрослые губки в основном сидячие животные, некоторые морские и пресноводные виды могут перемещаться по морскому дну со скоростью 1–4 мм (0,039–0,157 дюйма) в день в результате амебоподобных движений пинакоцитов и других клеток. Несколько видов могут сокращать все свое тело, а многие могут закрывать свои оскулы и остии. Молодь дрейфует или свободно плавает, в то время как взрослые остаются неподвижными. [18]

Губки не имеют отдельных кровеносных , дыхательных , пищеварительных и выделительных систем — вместо этого система потока воды поддерживает все эти функции. Они отфильтровывают частицы пищи из воды, протекающей через них. Частицы размером более 50 микрометров не могут попасть в отверстия, и пинакоциты потребляют их путем фагоцитоза (поглощение и внутриклеточное пищеварение). Частицы размером от 0,5 мкм до 50 мкм задерживаются в отверстиях, которые сужаются от внешнего к внутреннему концу. Эти частицы потребляются пинакоцитами или археоцитами , которые частично выдавливаются через стенки отверстий. Частицы размером с бактерии, менее 0,5 микрометров, проходят через отверстия и улавливаются и потребляются хоаноцитами . [18] Поскольку самые мелкие частицы являются наиболее распространенными, хоаноциты обычно захватывают 80% пищевых запасов губки. [29] Археоциты транспортируют пищу, упакованную в пузырьки, из клеток, которые напрямую переваривают пищу, в те, которые этого не делают. По крайней мере, один вид губки имеет внутренние волокна, которые функционируют как дорожки для использования археоцитами, переносящими питательные вещества, [18] и эти дорожки также перемещают инертные объекты. [20]
Раньше утверждалось, что стеклянные губки могут жить на питательных веществах, растворенных в морской воде, и очень не любят ил. [30] Однако исследование, проведенное в 2007 году, не нашло никаких доказательств этого и пришло к выводу, что они очень эффективно извлекают бактерии и другие микроорганизмы из воды (около 79%) и обрабатывают взвешенные частицы осадка, чтобы извлечь такую добычу. [31] Воротниковые тельца переваривают пищу и распределяют ее, завернув в пузырьки, которые транспортируются «моторными» молекулами динеина по пучкам микротрубочек , которые проходят по всему синцитию . [18]
Клетки губок поглощают кислород путем диффузии из воды в клетки, когда вода течет через тело, в которые также диффундируют углекислый газ и другие растворимые отходы, такие как аммиак . Археоциты удаляют минеральные частицы, которые угрожают заблокировать устья, транспортируют их через мезогил и обычно сбрасывают их в исходящий водный поток, хотя некоторые виды включают их в свои скелеты. [18]

В водах, где запас пищевых частиц очень скуден, некоторые виды охотятся на ракообразных и других мелких животных. На данный момент обнаружено всего 137 видов. [33] Большинство из них принадлежат к семейству Cladorhizidae , но несколько представителей Guitarridae и Esperiopsidae также являются плотоядными. [34] В большинстве случаев мало что известно о том, как они на самом деле захватывают добычу, хотя считается, что некоторые виды используют либо липкие нити, либо крючковатые спикулы . [34] [35] Большинство плотоядных губок обитают в глубоких водах, до 8840 м (5,49 миль), [36] и ожидается, что развитие методов исследования глубоководных районов океана приведет к открытию еще нескольких. [18] [34] Однако один вид был обнаружен в средиземноморских пещерах на глубине 17–23 м (56–75 футов), наряду с более обычными фильтрующими губками. Обитающие в пещерах хищники ловят ракообразных длиной менее 1 мм (0,039 дюйма), опутывая их тонкими нитями, переваривают их, обволакивая их дополнительными нитями в течение нескольких дней, а затем возвращаются к своей обычной форме; нет никаких доказательств того, что они используют яд . [36]
Большинство известных плотоядных губок полностью утратили систему тока воды и хоаноциты . Однако род Chondrocladia использует сильно модифицированную систему тока воды для надувания шарообразных структур, которые используются для захвата добычи. [34] [37]
Пресноводные губки часто содержат зеленые водоросли в качестве эндосимбионтов внутри археоцитов и других клеток и получают пользу от питательных веществ, вырабатываемых водорослями. Многие морские виды содержат другие фотосинтезирующие организмы, чаще всего цианобактерии , но в некоторых случаях динофлагелляты . Симбиотические цианобактерии могут составлять треть от общей массы живой ткани в некоторых губках, и некоторые губки получают от 48% до 80% своего энергетического обеспечения от этих микроорганизмов. [18] В 2008 году группа ученых из Университета Штутгарта сообщила, что спикулы, состоящие из кремния, проводят свет в мезогиль , где живут фотосинтезирующие эндосимбионты. [38] Губки, в которых обитают фотосинтезирующие организмы, чаще всего встречаются в водах с относительно скудными запасами пищевых частиц и часто имеют листовидную форму, которая максимизирует количество собираемого ими солнечного света. [20]
Недавно обнаруженная плотоядная губка, обитающая вблизи гидротермальных источников, является средой обитания бактерий , питающихся метаном , и переваривает некоторые из них. [20]
У губок нет сложной иммунной системы большинства других животных. Однако они отвергают трансплантаты от других видов, но принимают их от других членов своего собственного вида. У некоторых морских видов серые клетки играют ведущую роль в отторжении инородного материала. При вторжении они вырабатывают химическое вещество, которое останавливает движение других клеток в пораженной области, тем самым не давая нарушителю использовать внутренние транспортные системы губки. Если вторжение продолжается, серые клетки концентрируются в этой области и выделяют токсины, которые убивают все клетки в этой области. «Иммунная» система может оставаться в этом активированном состоянии до трех недель. [20]

Губки имеют три бесполых способа размножения: после фрагментации, почкованием и путем образования геммул . Фрагменты губок могут быть отделены течениями или волнами. Они используют подвижность своих пинакоцитов и хоаноцитов и изменение формы мезогила, чтобы снова прикрепиться к подходящей поверхности, а затем восстановить себя в виде небольших, но функциональных губок в течение нескольких дней. Те же возможности позволяют губкам, которые были выдавлены через тонкую ткань, регенерировать. [18] : 239 Фрагмент губки может регенерировать только в том случае, если он содержит как колленциты для производства мезогила , так и археоциты для производства всех других типов клеток. [29] Очень немногие виды размножаются почкованием. [18] : 90–94
Геммулы — это «стадии выживания», которые несколько морских губок и многие пресноводные виды производят тысячами, когда умирают, и которые некоторые, в основном пресноводные виды, регулярно производят осенью. Спонгоциты создают геммулы, обертывая оболочки спонгина, часто усиленные спикулами, вокруг скоплений археоцитов , которые полны питательных веществ. [18] : 87–88 Пресноводные геммулы могут также включать фотосинтезирующие симбионты. [39] Затем геммулы становятся спящими и в этом состоянии могут пережить холод, высыхание, недостаток кислорода и экстремальные колебания солености . [18] Пресноводные геммулы часто не оживают, пока температура не упадет, останется холодной в течение нескольких месяцев, а затем не достигнет почти «нормального» уровня. [39] Когда геммулы прорастают, археоциты вокруг внешней стороны кластера трансформируются в пинакоциты , мембрана над порой в оболочке лопается, кластер клеток медленно выходит наружу, и большинство оставшихся археоцитов трансформируются в другие типы клеток, необходимые для создания функционирующей губки. Геммулы одного вида, но разных особей могут объединить усилия, чтобы сформировать одну губку. [18] : 89–90 Некоторые геммулы сохраняются в родительской губке, и весной может быть трудно сказать, возродилась ли старая губка или была «реколонизирована» своими собственными геммулами. [39]
Большинство губок являются гермафродитами (функционируют как оба пола одновременно), хотя у губок нет гонад (репродуктивных органов). Сперма вырабатывается хоаноцитами или целыми камерами хоаноцитов, которые погружаются в мезогил и образуют сперматические кисты , в то время как яйца образуются путем трансформации археоцитов или хоаноцитов у некоторых видов. Каждое яйцо обычно приобретает желток , потребляя «клетки-кормилицы». Во время нереста сперматозоиды вырываются из своих цист и выбрасываются через оскулюм . Если они контактируют с другой губкой того же вида, поток воды переносит их к хоаноцитам, которые поглощают их, но вместо того, чтобы переварить их, метаморфозируют в амебоидную форму и переносят сперму через мезогил к яйцам, которые в большинстве случаев поглощают носителя и его груз. [18] : 77
Несколько видов выпускают оплодотворенные яйца в воду, но большинство сохраняют яйца до тех пор, пока они не вылупятся. Сохраняя яйца, родители могут передавать симбиотические микроорганизмы непосредственно своему потомству посредством вертикальной передачи , в то время как виды, которые выпускают свои яйца в воду, должны приобретать симбионтов горизонтально (комбинация обоих, вероятно, наиболее распространена, когда личинки с вертикально переданными симбионтами также приобретают других горизонтально). [40] [41] Существует четыре типа личинок, но все они представляют собой лецитотрофные (непитающиеся) шары клеток с внешним слоем клеток, жгутики или реснички которых позволяют личинкам двигаться. После плавания в течение нескольких дней личинки тонут и ползают, пока не найдут место для поселения. Большинство клеток трансформируются в археоциты, а затем в типы, соответствующие их местоположению в миниатюрной взрослой губке. [18] : 77 [42]
Эмбрионы стеклянной губки начинают с деления на отдельные клетки, но как только образуется 32 клетки, они быстро превращаются в личинок, которые снаружи имеют яйцевидную форму с полосой ресничек посередине, которую они используют для движения, но внутри имеют типичную для стеклянной губки структуру спикул с паутинообразной основной синцитией, обернутой вокруг и между ними, и хоаносинцитиями с несколькими воротничковыми тельцами в центре. Затем личинки покидают тела своих родителей. [43]
Цитологическая прогрессия оогенеза и сперматогенеза ( гаметогенеза ) у porifera очень похожа на таковую у других метазоа. [44] Большинство генов из классического набора мейотических генов, включая гены рекомбинации ДНК и репарации двухцепочечных разрывов, которые сохраняются у эукариот, экспрессируются у губок (например, Geodia hentscheli и Geodia phlegraei ). [44] Поскольку porifera считаются самыми ранними дивергентными животными, эти результаты указывают на то, что основной набор инструментов мейоза, включая возможности рекомбинации и репарации ДНК, присутствовал на ранних этапах эволюции эукариот. [44]
Губки в умеренных регионах живут максимум несколько лет, но некоторые тропические виды и, возможно, некоторые глубоководные виды могут жить 200 лет и более. Некоторые кальцинированные демо-губки растут всего на 0,2 мм (0,0079 дюйма) в год, и, если эта скорость постоянна, особи шириной 1 м (3,3 фута) должны быть около 5000 лет. Некоторые губки начинают половое размножение в возрасте всего нескольких недель, в то время как другие ждут, пока им не исполнится несколько лет. [18]
У взрослых губок отсутствуют нейроны или какой-либо другой вид нервной ткани . Однако большинство видов обладают способностью выполнять движения, которые координируются по всему телу, в основном сокращения пинакоцитов , сжимая водные каналы и, таким образом, вытесняя избыток осадка и другие вещества, которые могут вызывать закупорку. Некоторые виды могут сокращать оскулюм независимо от остальной части тела. Губки также могут сокращаться, чтобы уменьшить область, уязвимую для нападения хищников. В случаях, когда две губки слиты, например, если есть большая, но все еще неразделенная почка, эти волны сокращения медленно становятся координированными в обоих « сиамских близнецах ». Координирующий механизм неизвестен, но может включать химические вещества, похожие на нейротрансмиттеры . [45] Однако стеклянные губки быстро передают электрические импульсы через все части синцития и используют это, чтобы остановить движение своих жгутиков , если поступающая вода содержит токсины или избыточный осадок. [18] Считается, что миоциты отвечают за закрытие оскулюма и передачу сигналов между различными частями тела. [20]
Губки содержат гены, очень похожие на те, которые содержат «рецепт» для постсинаптической плотности , важной структуры приема сигнала в нейронах всех других животных. Однако у губок эти гены активируются только в «колбовых клетках», которые появляются только у личинок и могут обеспечивать некоторую сенсорную способность, пока личинки плавают. Это поднимает вопросы о том, представляют ли колбовые клетки предшественников настоящих нейронов или являются доказательством того, что предки губок имели настоящие нейроны, но потеряли их, когда приспособились к сидячему образу жизни. [46]
.jpg/1280px-Euplectella_aspergillum_(cropped).jpg)
Губки распространены по всему миру, обитая в самых разных океанических средах обитания, от полярных регионов до тропиков. [29] Большинство из них обитает в тихих, чистых водах, поскольку взмученный волнами или течениями осадок блокирует их поры, затрудняя им питание и дыхание. [30] Наибольшее количество губок обычно встречается на твердых поверхностях, таких как камни, но некоторые губки могут прикрепляться к мягкому осадку с помощью корневидного основания. [47]
Губки более многочисленны, но менее разнообразны в умеренных водах, чем в тропических водах, возможно, потому, что организмы, которые охотятся на губок, более многочисленны в тропических водах. [48] Стеклянные губки наиболее распространены в полярных водах и в глубинах умеренных и тропических морей, поскольку их очень пористая структура позволяет им извлекать пищу из этих бедных ресурсами вод с минимальными усилиями. Демоспонги и известковые губки многочисленны и разнообразны в более мелких неполярных водах. [49]
Различные классы губок обитают в разных средах обитания:
Губки с фотосинтезирующими эндосимбионтами производят в три раза больше кислорода , чем потребляют, а также больше органического вещества, чем потребляют. Такой вклад в ресурсы их среды обитания значителен вдоль Большого Барьерного рифа в Австралии , но относительно незначителен в Карибском море. [29]


Многие губки сбрасывают спикулы , образуя плотный ковер глубиной в несколько метров, который отпугивает иглокожих, которые в противном случае охотились бы на губки. [29] Они также вырабатывают токсины, которые не позволяют другим сидячим организмам, таким как мшанки или асцидии, расти на них или рядом с ними, что делает губки очень эффективными конкурентами за жизненное пространство. Одним из многих примеров является агелиферин .
Несколько видов, например, карибская огненная губка Tedania ignis , вызывают сильную сыпь у людей, которые с ними контактируют. [18] Черепахи и некоторые рыбы питаются в основном губками. Часто говорят, что губки вырабатывают химическую защиту от таких хищников. [18] Однако эксперименты не смогли установить связь между токсичностью химических веществ, вырабатываемых губками, и тем, как они кажутся рыбам на вкус, что снизило бы полезность химической защиты в качестве отпугивающих средств. Хищничество рыб может даже способствовать распространению губок путем отсоединения фрагментов. [20] Однако некоторые исследования показали, что рыбы отдают предпочтение губкам, не защищенным химически, [50] а другое исследование показало, что высокий уровень хищничества кораллов действительно предсказывал присутствие химически защищенных видов. [51]
Стеклянные губки не производят токсичных химикатов и живут в очень глубоких водах, где хищники встречаются редко. [30]
Spongillaflies, также известные как Spongillaflies ( Neuroptera , Sisyridae ), являются специализированными хищниками пресноводных губок. Самка откладывает яйца на растительность, нависающую над водой. Личинки вылупляются и падают в воду, где они ищут губки, чтобы питаться. Они используют свои удлиненные ротовые части, чтобы прокалывать губку и высасывать жидкость из нее. Личинки некоторых видов цепляются за поверхность губки, в то время как другие находят убежище во внутренних полостях губки. Полностью выросшие личинки покидают воду и плетут кокон, в котором окукливаются. [52]
Карибская губка Chondrilla nucula выделяет токсины, которые убивают коралловые полипы , позволяя губкам расти поверх коралловых скелетов. [18] Другие, особенно в семействе Clionaidae , используют едкие вещества, выделяемые их археоцитами, для прокладывания туннелей в скалах, кораллах и раковинах мертвых моллюсков . [18] Губки могут удалять до 1 м (3,3 фута) в год с рифов, создавая видимые выемки чуть ниже уровня отлива. [29]
Карибские губки рода Aplysina страдают от синдрома красных полос Aplysina. Это приводит к тому, что у Aplysina появляются одна или несколько полос ржавого цвета, иногда с прилегающими полосами некротической ткани. Эти поражения могут полностью окружать ветви губки. Болезнь, по-видимому, заразна и поражает примерно 10 процентов A. cauliformis на багамских рифах. [53] Полосы ржавого цвета вызваны цианобактерией , но неизвестно, вызывает ли этот организм заболевание на самом деле. [53] [54]
Помимо размещения фотосинтезирующих эндосимбионтов, [18] губки известны своим широким спектром сотрудничества с другими организмами. Относительно крупная инкрустирующая губка Lissodendoryx colombiensis чаще всего встречается на каменистых поверхностях, но расширила свой ареал на луга морской травы , позволяя себе быть окруженной или заросшей губками морской травы, которые неприятны местным морским звездам и, следовательно, защищают Lissodendoryx от них; в свою очередь, губки морской травы занимают более высокие позиции вдали от осадка морского дна. [55]
Креветки рода Synalpheus образуют колонии в губках, и каждый вид креветок обитает в разных видах губок, что делает Synalpheus одним из самых разнообразных родов ракообразных . В частности, Synalpheus regalis использует губку не только как источник пищи, но и как защиту от других креветок и хищников. [56] Около 16 000 особей обитают в одной губке логгерхед , питаясь более крупными частицами, которые собираются на губке, когда она фильтрует океан, чтобы прокормить себя. [57] Другие ракообразные, такие как раки-отшельники, обычно имеют особый вид губки, Pseudospongosorites , растущий на них, поскольку и губка, и краб занимают раковины брюхоногих моллюсков, пока краб и губка не перерастают раковину, в конечном итоге приводя к тому, что краб использует тело губки в качестве защиты вместо раковины, пока краб не найдет подходящую замену раковине. [58]

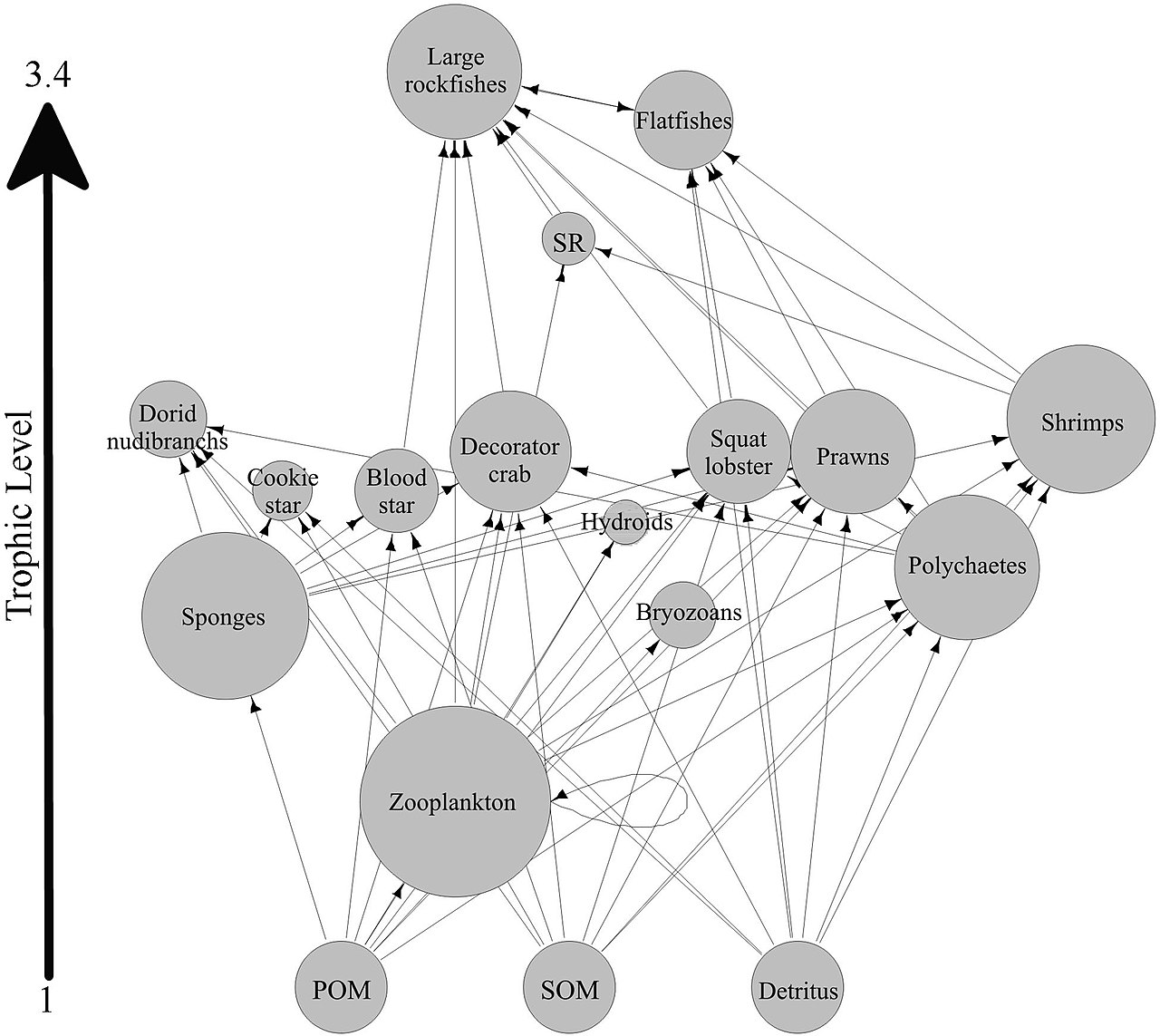
Большинство губок являются детритофагами , которые фильтруют органические частицы мусора и микроскопические формы жизни из океанской воды. В частности, губки играют важную роль детритофагов в пищевых сетях коралловых рифов, перерабатывая детрит на более высоких трофических уровнях . [61]
Была выдвинута гипотеза, что губки коралловых рифов облегчают передачу органического вещества, полученного из кораллов, их связанным с ними детритофагам посредством производства детрита губки, как показано на схеме. Несколько видов губок способны преобразовывать DOM, полученный из кораллов, в детрит губки, [62] [63] и переносить органическое вещество, произведенное кораллами, дальше по пищевой сети рифа. Кораллы выделяют органическое вещество как в виде растворенной, так и в виде частиц слизи, [64] [65] [66] [67], а также клеточный материал, такой как вытесненный Symbiodinium . [68] [69] [61]
Органическое вещество может передаваться от кораллов к губкам всеми этими путями, но DOM, вероятно, составляет наибольшую фракцию, так как большая часть (от 56 до 80%) коралловой слизи растворяется в толще воды, [65] а потеря кораллами связанного углерода из-за вытеснения Symbiodinium обычно незначительна (0,01%) [68] по сравнению с выделением слизи (до ~40%). [70] [71] Органическое вещество, полученное от кораллов, также может косвенно передаваться губкам через бактерии, которые также могут потреблять коралловую слизь. [72] [73] [74] [61]


Помимо симбиотических отношений один к одному , хозяин может стать симбиотиком с микробным консорциумом , что приведет к разнообразному микробиому губки . Губки способны содержать широкий спектр микробных сообществ , которые также могут быть очень специфичными. Микробные сообщества, которые образуют симбиотические отношения с губкой, могут составлять до 35% биомассы ее хозяина. [77] Термин для этих специфических симбиотических отношений, когда микробный консорциум образует пару с хозяином, называется голобиотической связью . Губка, а также микробное сообщество, связанное с ней, будут производить широкий спектр вторичных метаболитов , которые помогают защитить ее от хищников с помощью таких механизмов, как химическая защита . [78]
Некоторые из этих взаимоотношений включают эндосимбионтов внутри клеток бактериоцитов и цианобактерий или микроводорослей, обнаруженных под слоем клеток пинакодермы, где они способны получать наибольшее количество света, используемого для фототрофии. Они могут содержать более 50 различных микробных филумов и филумов-кандидатов, включая Alphaprotoebacteria, Actinomycetota , Chloroflexota , Nitrospirota , « Cyanobacteria », таксоны Gamma-, филум-кандидат Poribacteria и Thaumarchaea . [78]
Карл Линней , который классифицировал большинство видов сидячих животных как принадлежащих к отряду Zoophyta в классе Vermes , ошибочно определил род Spongia как растения в отряде Algae . [79] [ необходимо дальнейшее объяснение ] Долгое время после этого губки относили к подцарству Parazoa («рядом с животными»), отделенному от Eumetazoa , которые образовали остальную часть царства Animalia . [80]
Тип Porifera далее делится на классы , в основном в соответствии с составом их скелетов : [17] [29]
В 1970-х годах губки с массивными скелетами из карбоната кальция были отнесены к отдельному классу Sclerospongiae , также известному как «коралловые губки». [81] Однако в 1980-х годах было обнаружено, что все они являются представителями либо Calcarea, либо Demospongiae. [82]
На сегодняшний день научные публикации выявили около 9000 видов губок, [29] из которых: около 400 являются стеклянными губками; около 500 являются известковыми видами; а остальные являются демоспонгами. [18] Однако некоторые типы местообитаний, вертикальные скалы и стены пещер, а также галереи в скалах и коралловых валунах, были исследованы очень мало, даже в мелководных морях. [29]
Губки традиционно делились на три класса: известковые губки (Calcarea), стеклянные губки (Hexactinellida) и демоспонгии (Demospongiae). Однако исследования показали, что Homoscleromorpha , группа, которая, как считается, принадлежит к Demospongiae , имеет генетическую связь, хорошо отделенную от других классов губок. [13] : 153–154 Поэтому они недавно были признаны четвертым классом губок. [83] [84]
Губки делятся на классы в основном по составу их скелетов : [20] Они расположены в эволюционном порядке, как показано ниже, в порядке возрастания их эволюции сверху вниз:
Филогения губок активно обсуждалась с момента появления филогенетики . Первоначально считавшаяся наиболее базальным типом животных, теперь есть весомые доказательства того, что Ctenophora может носить этот титул. [86] [87] Кроме того, монофилия типа теперь находится под вопросом. Несколько исследований пришли к выводу, что все другие животные произошли от губок, и обычно восстанавливают, что известковые губки и Homoscleromorpha ближе к другим животным, чем к демо-губкам . [88] [89] Внутренние взаимоотношения Porifera оказались менее неопределенными. Тесная связь Homoscleromorpha и Calcarea была восстановлена почти во всех исследованиях, независимо от того, поддерживают ли они монофилию губок или эуметазоа. [88] [6] [84] [83] Положение стеклянных губок также довольно определенно, при этом большинство исследований восстанавливают их как сестру демо-губок. [83] [6] [88] Таким образом, неопределенность в основании генеалогического древа животных, вероятно, лучше всего представлена нижеприведенной кладограммой.


Хотя молекулярные часы и биомаркеры предполагают, что губки существовали задолго до кембрийского взрыва жизни, кремниевые спикулы, подобные спикулам демогубок, отсутствуют в палеонтологической летописи до кембрия. [90] Существует неподтвержденный отчет 2002 года о спикулах в породах, датируемых примерно 750 миллионами лет назад . [91] Хорошо сохранившиеся ископаемые губки, возраст которых составляет около 580 миллионов лет в эдиакарский период, были найдены в формации Доушаньто . [92] Эти окаменелости, которые включают: спикулы; пинакоциты ; пороциты ; археоциты ; склероциты ; и внутреннюю полость, были классифицированы как демогубки. Ископаемые остатки стеклянных губок были найдены примерно 540 миллионов лет назад в породах в Австралии, Китае и Монголии. [93] Ранние кембрийские губки из Мексики, принадлежащие к роду Kiwetinokia, демонстрируют свидетельства слияния нескольких более мелких спикул в одну большую спикулу. [94] Спикулы карбоната кальция известковых губок были обнаружены в раннекембрийских породах возрастом около 530–523 миллионов лет назад в Австралии. Другие вероятные демогубки были обнаружены в раннекембрийской фауне Чэнцзян возрастом от 525 до 520 миллионов лет назад . [95] Ископаемые останки, найденные на канадских северо-западных территориях, датируемые 890 миллионами лет назад, могут быть губками; если это открытие подтвердится, это говорит о том, что первые животные появились до неопротерозойского события оксигенации. [96]

Пресноводные губки, по-видимому, намного моложе, поскольку самые ранние известные окаменелости датируются средним эоценом , около 48–40 миллионов лет назад . [93] Хотя около 90% современных губок являются демогубками , окаменелые останки этого типа встречаются реже, чем останки других типов, поскольку их скелеты состоят из относительно мягкого спонгина, который плохо фоссилизуется. [97] Самые ранние симбионты губок известны с раннего силура . [98]
Химическим индикатором является 24-изопропилхолестан , который является стабильным производным 24-изопропилхолестерина , который, как говорят, вырабатывается демогубками , но не эуметазоа («истинными животными», т. е. книдариями и билатериями ). Поскольку хоанофлагелляты считаются ближайшими одноклеточными родственниками животных, группа ученых изучила биохимию и гены одного вида хоанофлагеллят . Они пришли к выводу, что этот вид не может вырабатывать 24-изопропилхолестерин, но что необходимо исследование более широкого спектра хоанофлагеллят, чтобы доказать, что ископаемый 24-изопропилхолестан мог вырабатываться только демогубками. [99] Хотя в предыдущей публикации сообщалось о следах химического 24-изопропилхолестана в древних породах, датируемых 1800 миллионами лет назад , [100] недавние исследования с использованием гораздо более точно датированной серии пород показали, что эти биомаркеры появляются только до конца оледенения Мариноа примерно 635 миллионов лет назад , [101] и что «анализ биомаркеров еще не выявил никаких убедительных доказательств того, что древние губки существовали до первого глобального обширного неопротерозойского ледникового эпизода (стертиан, ~ 713 миллионов лет назад в Омане)». Хотя утверждалось, что этот «губчатый биомаркер» мог произойти от морских водорослей, недавние исследования показывают, что способность водорослей вырабатывать этот биомаркер развилась только в карбоне ; как таковой, биомаркер по-прежнему решительно поддерживает присутствие демогубок в криогене. [102] [103] [104]
Археоциаты , которых некоторые классифицируют как тип коралловых губок, являются очень распространенными ископаемыми в породах раннего кембрия около 530–520 миллионов лет назад , но, по-видимому, вымерли к концу кембрия 490 миллионов лет назад . [95] Было высказано предположение, что они были созданы: губками; книдариями ; водорослями ; фораминиферами ; совершенно отдельным типом животных, Archaeocyatha; или даже совершенно отдельным царством жизни, обозначенным как Archaeata или Inferibionta. С 1990-х годов археоциаты рассматриваются как отличительная группа губок. [80]
Трудно вписать канцеллорид в классификации губок или более сложных животных. Анализ, проведенный в 1996 году, пришел к выводу, что они были тесно связаны с губками на том основании, что детальная структура склеритов канцеллорид («броневые пластины») похожа на структуру волокон спонгина, коллагенового белка , у современных кератозных (роговых) демо-губок, таких как Darwinella . [106] Однако другой анализ, проведенный в 2002 году, пришел к выводу, что канцеллориды не являются губками и могут занимать промежуточное положение между губками и более сложными животными, среди прочего, потому что их кожа была толще и более плотно соединена, чем у губок. [107] В 2008 году подробный анализ склеритов канцеллорид пришел к выводу, что они очень похожи на склериты халкиериид , подвижных билатеральных животных, которые выглядели как слизни в кольчуге и чьи окаменелости находят в породах от самого раннего кембрия до среднего кембрия. Если это верно, то это создаст дилемму, поскольку крайне маловероятно, что совершенно неродственные организмы могли бы развить такие похожие склериты независимо, но огромная разница в строении их тел затрудняет понимание того, как они могли быть тесно связаны. [105]

В 1990-х годах губки широко рассматривались как монофилетическая группа, все они произошли от общего предка , который сам был губкой, и как «сестринская группа» для всех других метазоа (многоклеточных животных), которые сами образуют монофилетическую группу. С другой стороны, некоторые анализы 1990-х годов также возродили идею о том, что ближайшими эволюционными родственниками животных являются хоанофлагелляты , одноклеточные организмы, очень похожие на хоаноциты губок , — что означало бы, что большинство метазоа произошли от очень губкоподобных предков и, следовательно, что губки могут не быть монофилетическими, поскольку одни и те же губкоподобные предки могли дать начало как современным губкам, так и негубкоподобным членам метазоа. [108]
Анализы с 2001 года пришли к выводу, что Eumetazoa (более сложные, чем губки) более тесно связаны с определенными группами губок, чем с другими группами губок. Такие выводы подразумевают, что губки не являются монофилетическими, потому что последний общий предок всех губок также был бы прямым предком Eumetazoa, которые не являются губками. Исследование 2001 года, основанное на сравнении рибосомной ДНК, пришло к выводу, что наиболее фундаментальное разделение внутри губок было между стеклянными губками и остальными, и что Eumetazoa более тесно связаны с известковыми губками (с карбонатными спикулами кальция), чем с другими типами губок. [108] В 2007 году один анализ, основанный на сравнении РНК , и другой, основанный в основном на сравнении спикул, пришли к выводу, что демо-губки и стеклянные губки более тесно связаны друг с другом, чем любая из них с известковыми губками, которые, в свою очередь, более тесно связаны с Eumetazoa. [93] [110]
Другие анатомические и биохимические данные связывают Eumetazoa с Homoscleromorpha , подгруппой demosponges. Сравнение в 2007 году ядерной ДНК , за исключением стеклянных губок и гребневиков , пришло к выводу, что:
Сперма Homoscleromorpha имеет общие черты со спермой Eumetazoa, которых нет у спермы других губок. Как у Homoscleromorpha, так и у Eumetazoa слои клеток связаны между собой путем прикрепления к коврообразной базальной мембране, состоящей в основном из коллагена «типа IV» , формы коллагена, не встречающейся в других губках, хотя волокна спонгина , которые укрепляют мезогил всех демогубок, похожи на коллаген «типа IV». [22]
Анализы, описанные выше, пришли к выводу, что губки являются наиболее близкими к предкам всех Metazoa, всех многоклеточных животных, включая как губки, так и более сложные группы. Однако другое сравнение в 2008 году 150 генов в каждом из 21 рода, от грибов до людей, но включая только два вида губок, предположило, что гребневики ( ctenophora ) являются наиболее базальной линией Metazoa, включенных в выборку. [111] [112] [113] [114] Если это верно, то либо современные гребневики развили свои сложные структуры независимо от других Metazoa, либо предки губок были более сложными, и все известные губки представляют собой радикально упрощенные формы. Исследование рекомендовало дальнейшие анализы с использованием более широкого спектра губок и других простых Metazoa, таких как Placozoa . [111]
Однако повторный анализ данных показал, что компьютерные алгоритмы, используемые для анализа, были введены в заблуждение из-за наличия специфических генов гребневиков, которые заметно отличались от генов других видов, в результате чего губки стали либо сестринской группой по отношению ко всем остальным животным, либо предковой парафилетической группой. [115] [116] «Генеалогические деревья», построенные с использованием комбинации всех доступных данных — морфологических, онтогенетических и молекулярных — пришли к выводу, что губки на самом деле являются монофилетической группой и вместе с книдариями образуют сестринскую группу по отношению к билатериям. [117] [118]
Очень большое и внутренне согласованное выравнивание 1719 белков в масштабе метазойных, опубликованное в 2017 году, показало, что (i) губки, представленные Homoscleromorpha, Calcarea, Hexactinellida и Demospongiae, являются монофилетическими, (ii) губки являются сестринской группой для всех других многоклеточных животных, (iii) гребневики появляются как вторая по раннему разветвлению линия животных, и (iv) плакозои появляются как третья линия животных, за которыми следуют книдарии, сестринские группы для билатерий . [119]
В марте 2021 года ученые из Дублина обнаружили дополнительные доказательства того, что губки являются сестринской группой для всех других животных [120] , а в мае 2023 года Шульц и др. обнаружили закономерности необратимых изменений в синтении генома, которые дают убедительные доказательства того, что гребневики являются сестринской группой для всех других животных [121] .

В отчете 1997 года описывалось использование бутылконосыми дельфинами губок в качестве инструмента в заливе Шарк в Западной Австралии. Дельфин прикрепляет морскую губку к своему роструму , который, предположительно, затем используется для защиты при поиске пищи на песчаном дне моря . [122] Поведение, известное как губкообразование , наблюдалось только в этом заливе и демонстрируется почти исключительно самками. Исследование, проведенное в 2005 году, пришло к выводу, что матери обучают этому поведению своих дочерей, и что все пользователи губок тесно связаны, что позволяет предположить, что это довольно недавнее нововведение. [16]


Карбонат кальция или кремниевые спикулы большинства родов губок делают их слишком грубыми для большинства видов использования, но два рода, Hippospongia и Spongia , имеют мягкие, полностью волокнистые скелеты. [13] : 88 Ранние европейцы использовали мягкие губки для многих целей, включая набивку для шлемов, переносную питьевую утварь и муниципальные фильтры для воды. До изобретения синтетических губок они использовались в качестве чистящих инструментов, аппликаторов для красок и керамической глазури и незаметных контрацептивов . Однако к середине 20-го века чрезмерный вылов рыбы привел как животных, так и отрасль к вымиранию . [123]
Многие объекты с губчатой текстурой теперь изготавливаются из веществ, не полученных из пориферанов. Синтетические губки включают в себя личные и бытовые чистящие средства , грудные имплантаты , [124] и контрацептивные губки . [125] Типичные используемые материалы - целлюлозная пена, полиуретановая пена и реже силиконовая пена.
«Губка» люффа, также называемая люфой , которая обычно продается для использования на кухне или в душе, получена не из животного, а в основном из волокнистого «скелета» губчатой тыквы ( Luffa aegyptiaca , Cucurbitaceae ). [126]
Губки обладают лечебным потенциалом из-за присутствия в них самих или в их микробных симбионтах химических веществ, которые могут быть использованы для борьбы с вирусами , бактериями , опухолями и грибками. [127] [128]

Не имея защитной оболочки или средств спасения, губки эволюционировали, чтобы синтезировать множество необычных соединений. Одним из таких классов являются окисленные производные жирных кислот, называемые оксилипинами . Было обнаружено, что члены этого семейства обладают противораковыми, антибактериальными и противогрибковыми свойствами. Один из примеров, выделенный из окинавских губок plakortis , plakoridine A , показал потенциал в качестве цитотоксина для клеток мышиной лимфомы. [129] [130]
{{cite journal}}: CS1 maint: неподходящий URL ( ссылка ){kind=link}
{kind=link}