Малая некодирующая молекула рибонуклеиновой кислоты
Pre-miRNA вместо Pri-miRNA в первой точке механизма. Схема действия микроРНК (miRNA) с мРНКПримеры петель микроРНК, где зрелые микроРНК показаны красным цветом
Микрорибонуклеиновая кислота ( микроРНК , miRNA , µRNA ) — это небольшие одноцепочечные некодирующие молекулы РНК, содержащие 21–23 нуклеотида . [1] Обнаруженные в растениях, животных и даже некоторых вирусах, miRNA участвуют в подавлении РНК и посттранскрипционной регуляции экспрессии генов . [2] [3] miRNA спариваются с комплементарными последовательностями в молекулах матричной РНК (мРНК), [4] затем подавляют указанные молекулы мРНК одним или несколькими из следующих процессов: [1] [5]
Расщепление цепи мРНК на две части.
Дестабилизация мРНК путем укорочения ее поли(А)-хвоста .
В клетках человека и других животных микроРНК в первую очередь действуют путем дестабилизации мРНК. [6] [7]
miRNAs напоминают малые интерферирующие РНК (siRNAs) пути РНК-интерференции (RNAi) , за исключением того, что miRNAs происходят из областей транскриптов РНК, которые складываются обратно, образуя короткие шпильки, тогда как siRNAs происходят из более длинных областей двухцепочечной РНК . [2] Геном человека может кодировать более 1900 miRNAs, [8] [9] Однако только около 500 человеческих miRNAs представляют собой настоящие miRNAs в вручную курируемой базе данных генов miRNA MirGeneDB . [10]
miRNAs широко распространены во многих типах клеток млекопитающих. [11] [12] Они, по-видимому, нацелены примерно на 60% генов человека и других млекопитающих. [13] [14] Многие miRNAs эволюционно консервативны, что подразумевает, что они выполняют важные биологические функции. [15] [1] Например, 90 семейств miRNAs были консервативны, по крайней мере, со времен общего предка млекопитающих и рыб, и большинство из этих консервативных miRNAs выполняют важные функции, как показали исследования, в которых гены одного или нескольких членов семейства были отключены у мышей. [1]
Первая miRNA была обнаружена в начале 1990-х годов. [19] Однако они не были признаны отдельным классом биологических регуляторов до начала 2000-х годов. [20] [21] [22] [23] [24] Исследования выявили различные наборы miRNA, экспрессируемых в различных типах клеток и тканях [12] [25], и множественные роли miRNA в развитии растений и животных, а также во многих других биологических процессах. [26] [27] [28] [29] [30] [31] [32] Аномальная экспрессия miRNA связана с болезненными состояниями. Терапия на основе miRNA находится в стадии изучения. [33] [34] [35] [36]
Первая miRNA была открыта в 1993 году группой под руководством Виктора Амброса , в которую входили Ли и Файнбаум. Однако для дополнительного понимания ее способа действия потребовалась одновременно опубликованная работа группы Гэри Рувкуна , в которую входили Уайтман и Ха. [19] [37] Эти группы опубликовали статьи один за другим о гене lin-4 , который, как известно, контролирует сроки развития личинок C. elegans, подавляя ген lin-14 . Когда Ли и др. выделили miRNA lin-4 , они обнаружили, что вместо того, чтобы производить мРНК, кодирующую белок, она производит короткие некодирующие РНК , одна из которых представляла собой РНК длиной ~22 нуклеотида, содержащую последовательности, частично комплементарные нескольким последовательностям в 3'-НТО мРНК lin-14 . [19] Было высказано предположение, что эта комплементарность ингибирует трансляцию мРНК lin-14 в белок LIN-14. В то время считалось, что малая РНК lin-4 является особенностью нематод .
В 2000 году была охарактеризована вторая малая РНК: let-7 РНК, которая подавляет lin-41, способствуя более позднему переходу в развитии у C. elegans . [20] Было обнаружено, что let -7 РНК сохраняется у многих видов, что привело к предположению, что let-7 РНК и дополнительные «малые временные РНК» могут регулировать сроки развития у различных животных, включая людей. [21]
Год спустя было обнаружено, что РНК lin-4 и let-7 являются частью большого класса малых РНК, присутствующих в клетках C. elegans , Drosophila и человека. [22] [23] [24] Многие РНК этого класса напоминали РНК lin-4 и let-7 , за исключением того, что их паттерны экспрессии обычно не соответствовали роли в регулировании сроков развития. Это предполагало, что большинство из них могут функционировать в других типах регуляторных путей. В этот момент исследователи начали использовать термин «микроРНК» для обозначения этого класса малых регуляторных РНК. [22] [23] [24]
Первым заболеванием человека, связанным с нарушением регуляции miRNA, был хронический лимфоцитарный лейкоз . При этом заболевании miRNA играют двойную роль, работая как супрессоры опухолей и онкогены. [38]
Номенклатура
В стандартной системе номенклатуры названия присваиваются экспериментально подтвержденным miRNA до публикации. [39] [40] За префиксом «miR» следует тире и число, последнее часто указывает порядок наименования. Например, miR-124 был назван и, вероятно, обнаружен до miR-456. Заглавная буква «miR-» относится к зрелой форме miRNA, в то время как строчная буква «mir-» относится к пре-miRNA и при-miRNA. [41] Гены, кодирующие miRNA, также называются с использованием того же трехбуквенного префикса в соответствии с соглашениями номенклатуры генов организмов. Например, официальные названия генов miRNA в некоторых организмах — « mir-1 у C. elegans и Drosophila, Mir1 у Rattus norvegicus и MIR25 у человека.
miRNA с почти идентичными последовательностями, за исключением одного или двух нуклеотидов, аннотируются дополнительной строчной буквой. Например, miR-124a тесно связана с miR-124b. Например:
hsa-miR-181a :aacauucaACgcugucggugAgu
hsa-miR-181b :aacauucaUUgcugucggugGgu
Пре-миРНК, при-миРНК и гены, которые приводят к 100% идентичным зрелым микроРНК, но которые расположены в разных местах генома, обозначены дополнительным суффиксом в виде тире-цифры. Например, пре-миРНК hsa-mir-194-1 и hsa-mir-194-2 приводят к идентичной зрелой микроРНК (hsa-miR-194), но происходят из генов, расположенных в разных областях генома.
Вид происхождения обозначается трехбуквенным префиксом, например, hsa-miR-124 — это человеческая ( Homo sapiens ) miRNA, а oar-miR-124 — это овечья ( Ovis aries ) miRNA. Другие распространенные префиксы включают «v» для вирусного (miRNA, кодируемого вирусным геномом) и «d» для miRNA Drosophila (плодовая мушка, обычно изучаемая в генетических исследованиях).
Когда две зрелые микроРНК происходят из противоположных плеч одной и той же пре-миРНК и встречаются примерно в одинаковых количествах, они обозначаются суффиксом -3p или -5p. (Раньше это различие также делалось с помощью «s» ( смысловой ) и «as» (антисмысловой)). Однако зрелая микроРНК, обнаруженная в одном плече шпильки, обычно гораздо более многочисленна, чем обнаруженная в другом плече, [2] в этом случае звездочка после названия указывает на зрелый вид, обнаруженный в низких количествах в противоположном плече шпильки. Например, miR-124 и miR-124* разделяют пре-миРНК шпильку, но в клетке обнаруживается гораздо больше miR-124.
Цели
Растительные miRNA обычно имеют почти идеальное спаривание с их мишенями мРНК, что вызывает репрессию генов посредством расщепления целевых транскриптов. [26] [42] Напротив, животные miRNA способны распознавать свои мишени мРНК, используя всего лишь 6–8 нуклеотидов (семенная область) на 5'-конце miRNA, [13] [43] [44] чего недостаточно для спаривания, чтобы вызвать расщепление целевых мРНК. [4] Комбинаторная регуляция является особенностью регуляции miRNA у животных. [4] [45] У данной miRNA могут быть сотни различных мишеней мРНК, и данная цель может регулироваться несколькими miRNA. [14] [46]
Оценки среднего числа уникальных матричных РНК, которые являются целями для репрессии типичной miRNA, различаются в зависимости от метода оценки, [47] но множественные подходы показывают, что miRNA млекопитающих могут иметь много уникальных целей. Например, анализ miRNA, высококонсервативных у позвоночных, показывает, что каждая имеет в среднем около 400 консервативных целей. [14] Аналогичным образом эксперименты показывают, что один вид miRNA может снизить стабильность сотен уникальных матричных РНК. [48] Другие эксперименты показывают, что один вид miRNA может подавлять производство сотен белков, но эта репрессия часто является относительно мягкой (гораздо менее чем в 2 раза). [49] [50]
Биогенез
До 40% генов miRNA могут находиться в интронах или даже экзонах других генов. [51] Обычно, хотя и не исключительно, они находятся в смысловой ориентации, [52] [53] и, таким образом, обычно регулируются вместе с их генами-хозяевами. [51] [54] [55]
Шаблон ДНК не является последним словом в производстве зрелой miRNA: 6% человеческих miRNA демонстрируют редактирование РНК ( IsomiRs ), сайт-специфическую модификацию последовательностей РНК для получения продуктов, отличных от тех, которые кодируются их ДНК. Это увеличивает разнообразие и масштаб действия miRNA за пределами того, что подразумевается только геномом.
Транскрипция
Гены miRNA обычно транскрибируются РНК-полимеразой II (Pol II). [56] [57] Полимераза часто связывается с промотором, находящимся рядом с последовательностью ДНК, кодируя то, что станет шпильковой петлей пре-miRNA. Полученный транскрипт закрывается специально модифицированным нуклеотидом на 5'-конце, полиаденилируется несколькими аденозинами (поли(А)-хвост), [56] [52] и сплайсируется . Животные miRNA изначально транскрибируются как часть одного плеча ~80-нуклеотидной РНК -петли , которая, в свою очередь, образует часть предшественника miRNA длиной в несколько сотен нуклеотидов, называемого pri-miRNA. [56] [52] Когда предшественник стебля-петли находится в 3'-UTR, транскрипт может служить pri-miRNA и мРНК. [52] РНК-полимераза III (Pol III) транскрибирует некоторые микроРНК, особенно те, которые содержат восходящие последовательности Alu , транспортные РНК (тРНК) и промоторы с широкими перемежающимися повторами млекопитающих (MWIR). [58]
Ядерная переработка
Кристаллическая структура человеческого белка Drosha в комплексе с C-концевыми спиралями двух молекул DGCR8 (зеленый). Drosha состоит из двух доменов рибонуклеазы III (синий и оранжевый); домена связывания двухцепочечной РНК (желтый); и домена соединителя/платформы (серый), содержащего два связанных иона цинка (сферы). Из PDB : 5B16 .
Одна pri-miRNA может содержать от одного до шести предшественников miRNA. Эти шпильковые петлевые структуры состоят примерно из 70 нуклеотидов каждая. Каждая шпилька окружена последовательностями, необходимыми для эффективной обработки.
Структура двухцепочечной РНК (dsRNA) шпилек в pri-miRNA распознается ядерным белком, известным как критический регион синдрома ДиДжорджи 8 (DGCR8 или «Pasha» у беспозвоночных ), названным так из-за его связи с синдромом ДиДжорджи . DGCR8 ассоциируется с ферментом Drosha , белком, который разрезает РНК, для формирования комплекса микропроцессора . [59] [60] В этом комплексе DGCR8 ориентирует каталитический домен РНКазы III Drosha для высвобождения шпилек из pri-miRNA путем расщепления РНК примерно на одиннадцать нуклеотидов от основания шпильки (одна спиральная dsRNA превращается в стебель). [61] [62] Полученный продукт имеет двухнуклеотидный выступ на своем 3'-конце; он имеет 3'-гидроксильную и 5'-фосфатную группы. Его часто называют пре-миРНК (предшественник-миРНК). Были идентифицированы мотивы последовательности ниже пре-миРНК, которые важны для эффективной обработки. [63] [64] [65]
Пре-миРНК, которые вырезаются непосредственно из интронов, минуя комплекс микропроцессора, известны как « миртроны ». [66] Миртроны были обнаружены у Drosophila , C. elegans и млекопитающих. [66] [67]
До 16% пре-миРНК могут быть изменены посредством редактирования ядерной РНК . [68] [69] [70] Чаще всего ферменты , известные как аденозиндезаминазы, действующие на РНК (ADAR), катализируют переходы аденозина в инозин (от A до I). Редактирование РНК может остановить ядерную обработку (например, pri-miR-142, что приводит к деградации рибонуклеазой Tudor-SN) и изменить последующие процессы, включая цитоплазматическую обработку miRNA и целевую специфичность (например, путем изменения затравочной области miR-376 в центральной нервной системе). [68]
Ядерный экспорт
Белок экспортина-5 человека (красный) в комплексе с Ran-GTP (желтый) и пре-микроРНК (зеленый), показывающий элемент распознавания с двумя выступающими нуклеотидами (оранжевый). Из PDB : 3A6P .
Шпильки пре-миРНК экспортируются из ядра в процессе, в котором участвует нуклеоцитоплазматический челнок экспортин-5 . Этот белок, член семейства кариоферинов , распознает двухнуклеотидный выступ, оставленный ферментом РНКазы III Дроша на 3'-конце шпильки пре-миРНК. Транспорт, опосредованный экспортином-5, в цитоплазму зависит от энергии и использует гуанозинтрифосфат (ГТФ), связанный с белком Ran . [71]
Цитоплазматическая обработка
В цитоплазме шпилька pre-miRNA расщепляется ферментом РНКазой III Dicer . [72] Эта эндорибонуклеаза взаимодействует с 5' и 3' концами шпильки [73] и разрезает петлю, соединяющую 3' и 5' плечи, в результате чего получается несовершенный дуплекс miRNA:miRNA* длиной около 22 нуклеотидов. [72] Общая длина шпильки и размер петли влияют на эффективность обработки Dicer. Несовершенная природа спаривания miRNA:miRNA* также влияет на расщепление. [72] [74] Некоторые из богатых G pre-miRNA потенциально могут принимать структуру G-квадруплекса в качестве альтернативы канонической структуре стебель-петля. Например, человеческая pre-miRNA 92b принимает структуру G-квадруплекса , которая устойчива к расщеплению, опосредованному Dicer, в цитоплазме . [75] Хотя любая из цепей дуплекса может потенциально действовать как функциональная микроРНК, только одна цепь обычно включается в комплекс подавления экспрессии, индуцированный РНК (RISC), где взаимодействуют микроРНК и ее целевая мРНК.
Хотя большинство miRNAs находятся внутри клетки, некоторые miRNAs, обычно известные как циркулирующие miRNAs или внеклеточные miRNAs, также были обнаружены во внеклеточной среде, включая различные биологические жидкости и среды для культивирования клеток. [76] [77]
Биогенез в растениях
Биогенез miRNA в растениях отличается от биогенеза животных в основном этапами ядерного процессинга и экспорта. Вместо того, чтобы быть расщепленным двумя разными ферментами, один раз внутри и один раз снаружи ядра, оба расщепления растительной miRNA выполняются гомологом Dicer, называемым Dicer-like1 (DL1). DL1 экспрессируется только в ядре растительных клеток, что указывает на то, что обе реакции происходят внутри ядра. Перед тем, как дуплексы miRNA:miRNA* растений транспортируются из ядра, их 3'-выступы метилируются белком РНК-метилтрансферазы, называемым Hua-Enhancer1 (HEN1). Затем дуплекс транспортируется из ядра в цитоплазму белком, называемым Hasty (HST), гомологом Exportin 5, где они разбираются, и зрелая miRNA включается в RISC. [78]
РНК-индуцированный комплекс подавления
Зрелая miRNA является частью активного комплекса подавления генов, индуцированного РНК (RISC), содержащего Dicer и множество связанных белков. [79] RISC также известен как комплекс микроРНК-рибонуклеопротеинов (miRNP); [80] RISC с включенной miRNA иногда называют «miRISC».
Считается, что обработка пре-miRNA с помощью Dicer сопряжена с раскручиванием дуплекса. Обычно в miRISC включается только одна нить, выбранная на основе ее термодинамической нестабильности и более слабого спаривания оснований на 5'-конце по сравнению с другой нитью. [81] [82] [83] Положение стебель-петли также может влиять на выбор нити. [84] Другая нить, называемая пассажирской нитью из-за ее более низких уровней в устойчивом состоянии, обозначена звездочкой (*) и обычно деградирует. В некоторых случаях обе нити дуплекса жизнеспособны и становятся функциональными miRNA, которые нацелены на различные популяции мРНК. [85]
AGO2 (серый) в комплексе с микроРНК (светло-голубой) и ее целевой мРНК (темно-синий)
Члены семейства белков Argonaute (Ago) играют центральную роль в функции RISC. Аргонавты необходимы для индуцированного miRNA сайленсинга и содержат два консервативных домена связывания РНК: домен PAZ, который может связывать одноцепочечный 3'-конец зрелой miRNA, и домен PIWI , который структурно напоминает рибонуклеазу-H и функционирует для взаимодействия с 5'-концом направляющей цепи. Они связывают зрелую miRNA и ориентируют ее для взаимодействия с целевой мРНК. Некоторые аргонавты, например, человеческий Ago2, напрямую расщепляют целевые транскрипты; аргонавты также могут привлекать дополнительные белки для достижения трансляционной репрессии. [86] Геном человека кодирует восемь белков аргонавтов, разделенных по сходству последовательностей на два семейства: AGO (четыре члена присутствуют во всех клетках млекопитающих и называются E1F2C/hAgo у людей) и PIWI (обнаружен в зародышевых и гемопоэтических стволовых клетках). [80] [86]
Подавление гена может происходить либо через деградацию мРНК, либо через предотвращение трансляции мРНК. Например, miR16 содержит последовательность, комплементарную богатому AU элементу [90] , обнаруженному в 3'UTR многих нестабильных мРНК, таких как TNF альфа или GM-CSF . [91] Было продемонстрировано, что при полной комплементарности между последовательностью мРНК и целевой мРНК Ago2 может расщеплять мРНК и приводить к прямой деградации мРНК. При отсутствии комплементарности подавление достигается путем предотвращения трансляции. [48] Связь мРНК и ее целевой мРНК может быть основана на простой отрицательной регуляции целевой мРНК, но, по-видимому, общим сценарием является использование «когерентной петли прямой связи », «взаимной петли отрицательной обратной связи» (также называемой двойной отрицательной петлей) и «петли положительной обратной связи/прямой связи». Некоторые мРНК работают как буферы случайных изменений экспрессии генов, возникающих из-за стохастических событий в транскрипции, трансляции и стабильности белка. Такая регуляция обычно достигается за счет действия циклов отрицательной обратной связи или некогерентных циклов прямой связи, разобщающих выход белка из транскрипции мРНК.
Оборот
Оборот зрелой miRNA необходим для быстрых изменений в профилях экспрессии miRNA. Во время созревания miRNA в цитоплазме поглощение белком Argonaute, как полагают, стабилизирует направляющую цепь, в то время как противоположная (* или «пассажирская») цепь преимущественно разрушается. В так называемой стратегии «Используй или потеряй», Argonaute может преимущественно сохранять miRNA со многими целями, а не miRNA с малым количеством или без целей, что приводит к деградации нецелевых молекул. [92]
Распад зрелых miRNA в Caenorhabditis elegans опосредован 5'-3' экзорибонуклеазой XRN2 , также известной как Rat1p. [93] У растений члены семейства SDN (малые РНК-деградирующие нуклеазы) разрушают miRNA в противоположном направлении (3'-5'). Аналогичные ферменты закодированы в геномах животных, но их роль не описана. [92]
Несколько модификаций miRNA влияют на стабильность miRNA. Как показали исследования на модельном организме Arabidopsis thaliana (кресс-салат Таля), зрелые растительные miRNA, по-видимому, стабилизируются добавлением метильных групп на 3'-конце. 2'-O-сопряженные метильные группы блокируют добавление остатков урацила (U) ферментами уридилтрансфераз , модификацию, которая может быть связана с деградацией miRNA. Однако уридилирование может также защищать некоторые miRNA; последствия этой модификации не полностью изучены. Сообщалось об уридилировании некоторых животных miRNA. Как растительные, так и животные miRNA могут быть изменены добавлением остатков аденина (A) к 3'-концу miRNA. Дополнительный A, добавленный к концу miR-122 млекопитающих , обогащенной печенью miRNA, важной при гепатите C , стабилизирует молекулу, а растительные miRNA, заканчивающиеся остатком аденина, имеют более медленные скорости распада. [92]
Клеточные функции
Взаимодействие микроРНК с процессом трансляции белка. Показаны несколько механизмов репрессии трансляции: M1) в процессе инициации, предотвращая сборку комплекса инициации или привлекая рибосомальную субъединицу 40S; M2) в сборке рибосомы; M3) в процессе трансляции; M7, M8) в процессе деградации мРНК. [94] 40S и 60S являются легкими и тяжелыми компонентами рибосомы, 80S является собранной рибосомой, связанной с мРНК, eIF4F является фактором инициации трансляции, PABC1 является связывающим белком Poly-A, а «cap» является структурой кэпа мРНК, необходимой для кольцевания мРНК (которая может быть нормальной m7G-cap или модифицированной A-cap). Инициация мРНК может происходить независимо от кэпа, посредством привлечения 40S к IRES ( внутренний сайт входа рибосомы ), расположенному в области 5'UTR. Фактическая работа по подавлению РНК выполняется RISC, в котором основной каталитической субъединицей является один из белков-аргонавтов (AGO), а miRNA служит шаблоном для распознавания определенных последовательностей мРНК.
Функция miRNA, по-видимому, заключается в регуляции генов. Для этой цели miRNA комплементарна части одной или нескольких матричных РНК (мРНК). Животные miRNA обычно комплементарны участку в 3'-НТО , тогда как растительные miRNA обычно комплементарны кодирующим областям мРНК. [95] Идеальное или почти идеальное спаривание оснований с целевой РНК способствует расщеплению РНК. [96] Это основной режим растительных miRNA. [97] У животных совпадения несовершенны.
Для того, чтобы частично комплементарные микроРНК распознавали свои цели, нуклеотиды 2–7 miRNA (ее «семенной регион» [13] [43] ) должны быть идеально комплементарными. [98] Животные miRNA ингибируют трансляцию белка целевой мРНК [99] (это присутствует, но менее распространено у растений). [97] Частично комплементарные микроРНК также могут ускорять деаденилирование , вызывая более раннюю деградацию мРНК. [100] Хотя деградация мРНК, нацеленной на miRNA, хорошо документирована, вопрос о том, достигается ли трансляционная репрессия посредством деградации мРНК, трансляционного ингибирования или комбинации этих двух, является предметом горячих споров. Недавние исследования miR-430 у данио-рерио, а также по бантам-miRNA и miR-9 в культивируемых клетках дрозофилы , показывают, что трансляционная репрессия вызвана нарушением инициации трансляции , независимо от деаденилирования мРНК. [101] [102]
Ингибирование транскрипции посредством реорганизации хроматина, опосредованной микроРНК, с последующим подавлением генов.
Часто невозможно различить эти механизмы, используя экспериментальные данные о стационарных скоростях реакции. Тем не менее, они дифференцированы в динамике и имеют различные кинетические сигнатуры . [94]
В отличие от растительных микроРНК, животные микроРНК нацелены на различные гены. [43] Однако гены, участвующие в функциях, общих для всех клеток, таких как экспрессия генов, имеют относительно меньше целевых участков микроРНК и, по-видимому, находятся под отбором, чтобы избежать нацеливания микроРНК. [105] Существует сильная корреляция между регуляцией гена ITPR и mir-92 и mir-19. [106]
dsRNA также может активировать экспрессию генов , механизм, который был назван «малой РНК-индуцированной активацией генов» или РНКа . dsRNA, нацеленные на промоторы генов, могут вызывать мощную транскрипционную активацию связанных генов. Это было продемонстрировано в клетках человека с использованием синтетических dsRNA, называемых малыми активирующими РНК (saRNAs), [107] , но также было продемонстрировано для эндогенных микроРНК. [108]
Взаимодействия между микроРНК и комплементарными последовательностями на генах и даже псевдогенами , которые разделяют гомологию последовательностей, считаются обратным каналом связи, регулирующим уровни экспрессии между паралогичными генами (генами, имеющими схожую структуру, указывающую на расхождение с общим предковым геном). Получив название «конкурирующие эндогенные РНК» ( ceRNAs ), эти микроРНК связываются с «элементами ответа микроРНК» на генах и псевдогенах и могут дать еще одно объяснение персистенции некодирующей ДНК . [109]
miRNA также обнаруживаются как внеклеточные циркулирующие miRNA . [110] Циркулирующие miRNA высвобождаются в биологические жидкости, включая кровь и спинномозговую жидкость , и могут быть доступны в качестве биомаркеров при ряде заболеваний. [110] [111] Некоторые исследования показывают, что груз мРНК экзосом может играть роль в имплантации, они могут разрушать адгезию между трофобластом и эндометрием или поддерживать адгезию путем подавления или повышения экспрессии генов, участвующих в адгезии/инвазии. [112]
Более того, miRNA, как miR-183/96/182, по-видимому, играет ключевую роль в циркадном ритме . [113]
Эволюция
miRNAs хорошо сохраняются как у растений, так и у животных и считаются жизненно важным и эволюционно древним компонентом регуляции генов. [114] [115] [116] [117] [118] В то время как основные компоненты пути микроРНК сохраняются у растений и животных , репертуары miRNA в двух царствах, по-видимому, возникли независимо с различными основными способами действия. [119] [120]
МикроРНК являются полезными филогенетическими маркерами из-за их, по-видимому, низкой скорости эволюции. [121] Происхождение микроРНК как регуляторного механизма, развившегося из предыдущего механизма РНК-интерференции, который изначально использовался в качестве защиты от экзогенного генетического материала, такого как вирусы. [122] Их происхождение, возможно, позволило развить морфологические инновации и, сделав экспрессию генов более специфичной и «тонко настраиваемой», позволило зародиться сложным органам [123] и, возможно, в конечном итоге, сложной жизни. [118] Быстрые всплески морфологических инноваций, как правило, связаны с высокой скоростью накопления микроРНК. [121] [123]
Новые микроРНК создаются несколькими способами. Новые микроРНК могут возникать из случайного образования шпилек в «некодирующих» участках ДНК (т. е. интронах или межгенных областях), а также путем дупликации и модификации существующих микроРНК. [124] МикроРНК также могут образовываться из инвертированных дупликаций белок-кодирующих последовательностей, что позволяет создавать структуру шпильки с загнутыми концами. [125] Скорость эволюции (т. е. замены нуклеотидов) в недавно возникших микроРНК сопоставима с таковой в других местах в некодирующей ДНК, что подразумевает эволюцию путем нейтрального дрейфа; однако более старые микроРНК имеют гораздо более низкую скорость изменений (часто менее одной замены за сто миллионов лет), [118] что предполагает, что как только микроРНК приобретает функцию, она подвергается очищающему отбору. [124] Отдельные регионы внутри гена miRNA сталкиваются с различным эволюционным давлением, где регионы, которые жизненно важны для обработки и функционирования, имеют более высокий уровень консервации. [126] На этом этапе микроРНК редко теряется из генома животного, [118] хотя более новые микроРНК (таким образом, предположительно нефункциональные) часто теряются. [124] У Arabidopsis thaliana чистый поток генов микроРНК, как было предсказано, составляет от 1,2 до 3,3 генов за миллион лет. [127] Это делает их ценным филогенетическим маркером, и они рассматриваются как возможное решение нерешенных филогенетических проблем, таких как взаимоотношения членистоногих . [128] С другой стороны, во многих случаях микроРНК плохо коррелируют с филогенией, и возможно, что их филогенетическое соответствие в значительной степени отражает ограниченную выборку микроРНК. [129]
МикроРНК присутствуют в геномах большинства эукариотических организмов, от бурых водорослей [130] до животных. Однако разница в том, как функционируют эти микроРНК и как они обрабатываются, предполагает, что микроРНК возникли независимо у растений и животных. [131]
Если сосредоточиться на животных, то в геноме Mnemiopsis leidyi [132] по-видимому отсутствуют узнаваемые микроРНК, а также ядерные белки Drosha и Pasha , которые имеют решающее значение для канонического биогенеза микроРНК. Это единственное животное, о котором до сих пор сообщалось, что у него отсутствует Drosha. МикроРНК играют важную роль в регуляции экспрессии генов у всех не-гребневиков, исследованных до сих пор, за исключением Trichoplax adhaerens , первого известного представителя типа Placozoa . [133]
К марту 2010 года у всех видов было идентифицировано более 5000 различных микроРНК. [134] В то время как у бактерий встречаются короткие последовательности РНК (от 50 до сотен пар оснований) с примерно сопоставимой функцией, у бактерий отсутствуют настоящие микроРНК. [135]
Экспериментальное обнаружение и манипуляция
В то время как исследователи сосредоточились на экспрессии miRNA в физиологических и патологических процессах, появились различные технические переменные, связанные с изоляцией микроРНК. Стабильность хранимых образцов miRNA была поставлена под сомнение. [77] микроРНК деградируют гораздо легче, чем мРНК, отчасти из-за их длины, но также из-за повсеместно присутствующих РНКаз . Это делает необходимым охлаждение образцов на льду и использование оборудования, свободного от РНКаз . [136]
Экспрессию микроРНК можно количественно оценить в двухэтапном процессе полимеразной цепной реакции модифицированной ОТ-ПЦР с последующей количественной ПЦР . Вариации этого метода позволяют достичь абсолютной или относительной количественной оценки. [137] микроРНК также можно гибридизировать с микроматрицами , слайдами или чипами с зондами для сотен или тысяч мишеней микроРНК, так что относительные уровни микроРНК можно определить в разных образцах. [138] микроРНК можно как обнаружить, так и профилировать с помощью методов высокопроизводительного секвенирования ( секвенирование микроРНК ). [139] Активность микроРНК можно экспериментально ингибировать с помощью олигонуклеотида с заблокированной нуклеиновой кислотой (LNA) , олигонуклеотида морфолино [140] [141] или олигонуклеотида 2'-O-метил РНК. [142] Конкретную микроРНК можно заглушить с помощью комплементарного антагомира . Созревание микроРНК можно ингибировать в нескольких точках с помощью стерически блокирующих олигонуклеотидов. [143] [144] Целевой сайт miRNA транскрипта mRNA также может быть заблокирован стерически блокирующим олиго. [145] Для обнаружения miRNA "in situ" можно использовать зонды LNA [146] или Morpholino [147] . Закрытая конформация LNA приводит к улучшенным свойствам гибридизации и повышает чувствительность и селективность, что делает его идеальным для обнаружения коротких miRNA. [148]
Высокопроизводительная количественная оценка miRNA подвержена ошибкам из-за большей дисперсии (по сравнению с мРНК ), которая сопровождается методологическими проблемами. Поэтому экспрессия мРНК часто анализируется для проверки эффектов miRNA на их уровнях (например, в [149] ). Базы данных могут использоваться для сопряжения данных мРНК и miRNA, которые предсказывают цели miRNA на основе их базовой последовательности. [150] [151] Хотя это обычно делается после обнаружения интересующих miRNA (например, из-за высоких уровней экспрессии), были предложены идеи для инструментов анализа, которые интегрируют информацию об экспрессии мРНК и miRNA. [152] [153]
Болезни людей и животных
Так же, как miRNA участвует в нормальном функционировании эукариотических клеток, так и нарушение регуляции miRNA связано с заболеванием. Вручную курируемая, общедоступная база данных miR2Disease документирует известные связи между нарушением регуляции miRNA и заболеванием человека. [154]
Наследственные заболевания
Мутация в области семени miR-96 вызывает наследственную прогрессирующую потерю слуха. [155]
Мутация в области семени miR-184 вызывает наследственный кератоконус с передней полярной катарактой. [156]
Удаление кластера miR-17~92 вызывает дефекты скелета и роста. [157]
Рак
Роль miRNA в раковой клетке
Первым заболеванием человека, которое, как известно, было связано с нарушением регуляции miRNA, был хронический лимфоцитарный лейкоз . [158] Многие другие miRNA также связаны с раком и, соответственно, иногда называются « онкомирами ». [159] В злокачественных B-клетках miRNA участвуют в путях, фундаментальных для развития B-клеток, таких как сигнализация B-клеточного рецептора (BCR), миграция/адгезия B-клеток, межклеточные взаимодействия в иммунных нишах и выработка и переключение классов иммуноглобулинов. MiRNA влияют на созревание B-клеток, генерацию пре-, маргинальной зоны, фолликулярных, B1, плазменных и памяти B-клеток. [160]
Другая роль miRNA в раке заключается в использовании уровня их экспрессии для прогнозирования. В образцах NSCLC низкие уровни miR-324 a могут служить индикатором плохой выживаемости. [161] Либо высокие уровни miR-185, либо низкие уровни miR-133b могут коррелировать с метастазами и плохой выживаемостью при колоректальном раке . [162]
Более того, определенные miRNA могут быть связаны с определенными гистологическими подтипами колоректального рака. Например, было показано, что уровни экспрессии miR-205 и miR-373 увеличиваются при муцинозном колоректальном раке и муцин-продуцирующем язвенном колите, ассоциированном с раком толстой кишки, но не при спорадической аденокарциноме толстой кишки, в которой отсутствуют муцинозные компоненты. [163] Исследования in vitro показали, что miR-205 и miR-373 могут функционально вызывать различные особенности муцинозно-ассоциированной неопластической прогрессии в эпителиальных клетках кишечника. [163]
Пролиферация клеток гепатоцеллюлярной карциномы может возникать из-за взаимодействия miR-21 с MAP2K3, геном-репрессором опухоли. [164] Оптимальное лечение рака включает в себя точное выявление пациентов для терапии стратифицированного риска. Те, у кого быстрый ответ на первоначальное лечение, могут выиграть от укороченных схем лечения, что показывает ценность точных измерений ответа на заболевание. Бесклеточные циркулирующие miRNA (cimiRNA) очень стабильны в крови, сверхэкспрессируются при раке и поддаются количественной оценке в диагностической лаборатории. При классической лимфоме Ходжкина плазменные miR-21, miR-494 и miR-1973 являются перспективными биомаркерами ответа на заболевание. [165] Циркулирующие miRNA могут помочь в принятии клинических решений и интерпретации позитронно-эмиссионной томографии в сочетании с компьютерной томографией . Их можно проводить на каждой консультации для оценки ответа на заболевание и выявления рецидива.
МикроРНК могут быть потенциально использованы в качестве инструментов или мишеней для лечения различных видов рака. [166] В нескольких исследованиях было обнаружено, что конкретная микроРНК, miR-506, действует как антагонист опухоли. Было обнаружено, что значительное количество образцов рака шейки матки имеют пониженную экспрессию miR-506. Кроме того, miR-506 работает над стимулированием апоптоза клеток рака шейки матки через свой прямой целевой фактор транскрипции пути hedgehog, Gli3. [167] [168]
Восстановление ДНК и рак
Многие микроРНК могут напрямую воздействовать на гены клеточного цикла и ингибировать их , чтобы контролировать пролиферацию клеток . Новая стратегия лечения опухолей заключается в ингибировании пролиферации опухолевых клеток путем восстановления дефектного пути микроРНК в опухолях. [169]
Рак вызывается накоплением мутаций либо из-за повреждения ДНК, либо из-за неисправленных ошибок в репликации ДНК . [170] Дефекты в репарации ДНК вызывают накопление мутаций, что может привести к раку. [171] Несколько генов, участвующих в репарации ДНК, регулируются микроРНК. [172]
Мутации зародышевой линии в генах репарации ДНК вызывают только 2–5% случаев рака толстой кишки . [173] Однако измененная экспрессия микроРНК, вызывающая дефицит репарации ДНК, часто связана с раком и может быть важным причинным фактором. Среди 68 спорадических случаев рака толстой кишки со сниженной экспрессией белка репарации несоответствий ДНК MLH1 большинство оказались дефицитными из-за эпигенетического метилирования острова CpG гена MLH1 . [174] Однако до 15% дефицитов MLH1 при спорадических случаях рака толстой кишки, по-видимому, были вызваны чрезмерной экспрессией микроРНК miR-155, которая подавляет экспрессию MLH1. [175]
В 29–66% [176] [177] глиобластом репарация ДНК является недостаточной из-за эпигенетического метилирования гена MGMT , что снижает экспрессию белка MGMT. Однако в 28% глиобластом белок MGMT является дефицитным, но промотор MGMT не метилирован. [ 176] В глиобластомах без метилированных промоторов MGMT уровень микроРНК miR-181d обратно коррелирует с экспрессией белка MGMT, а прямой целью miR-181d является мРНК MGMT 3'UTR ( три основных нетранслируемых региона мРНК MGMT). [176] Таким образом, в 28% глиобластом повышенная экспрессия miR-181d и сниженная экспрессия фермента репарации ДНК MGMT могут быть причинным фактором.
Белки HMGA (HMGA1a, HMGA1b и HMGA2) участвуют в развитии рака, и экспрессия этих белков регулируется микроРНК. Экспрессия HMGA практически не обнаруживается в дифференцированных тканях взрослых, но повышена при многих видах рака. Белки HMGA представляют собой полипептиды из ~100 аминокислотных остатков, характеризующиеся модульной организацией последовательностей. Эти белки имеют три высокоположительно заряженных региона, называемых крючками AT , которые связывают малую бороздку участков ДНК, богатых AT, в определенных регионах ДНК. Человеческие новообразования, включая карциномы щитовидной железы, предстательной железы, шейки матки, прямой и толстой кишки, поджелудочной железы и яичников, показывают сильное увеличение белков HMGA1a и HMGA1b. [178] Трансгенные мыши с HMGA1, нацеленным на лимфоидные клетки, развивают агрессивную лимфому, показывая, что высокая экспрессия HMGA1 связана с раком и что HMGA1 может действовать как онкоген. [179] Белок HMGA2 специфически воздействует на промотор ERCC1 , тем самым снижая экспрессию этого гена репарации ДНК. [180] Экспрессия белка ERCC1 была недостаточной в 100% из 47 оцененных случаев рака толстой кишки (хотя степень участия HGMA2 неизвестна). [181]
Однонуклеотидные полиморфизмы (SNP) могут изменять связывание miRNA на 3'UTR, например, в случае hsa-mir181a и hsa-mir181b на гене-супрессоре опухолей CDON. [182]
Сердечное заболевание
Глобальная роль функции miRNA в сердце была исследована путем условного ингибирования созревания miRNA в мышином сердце. Это показало, что miRNA играют существенную роль в его развитии. [183] [184] Исследования по профилированию экспрессии miRNA показывают, что уровни экспрессии определенных miRNA изменяются в больных сердцах человека, указывая на их участие в кардиомиопатиях . [185] [186] [187] Кроме того, исследования животных на определенных miRNA выявили различные роли miRNA как во время развития сердца, так и в патологических состояниях, включая регуляцию ключевых факторов, важных для кардиогенеза, реакции гипертрофического роста и сердечной проводимости. [184] [188] [189] [190] [191] [192] Другая роль miRNA при сердечно-сосудистых заболеваниях заключается в использовании их уровней экспрессии для диагностики, прогнозирования или стратификации риска. [193] miRNA в моделях животных также были связаны с метаболизмом и регуляцией холестерина.
миРНК-712
Мышиная микроРНК-712 является потенциальным биомаркером (т. е. предиктором) атеросклероза , сердечно-сосудистого заболевания артериальной стенки, связанного с задержкой липидов и воспалением. [194] Неламинарный кровоток также коррелирует с развитием атеросклероза, поскольку механосенсоры эндотелиальных клеток реагируют на силу сдвига нарушенного потока (d-flow). [195] Ряд проатерогенных генов, включая матриксные металлопротеиназы (ММП), активируются d-flow, [195] опосредуя провоспалительные и проангиогенные сигналы. Эти результаты были обнаружены в лигированных сонных артериях мышей, чтобы имитировать эффекты d-flow. В течение 24 часов ранее существовавшая незрелая miR-712 сформировала зрелую miR-712, что позволяет предположить, что miR-712 чувствительна к потоку. [195] В соответствии с этими результатами, miR-712 также повышается в эндотелиальных клетках, подвергающихся естественному d-потоку в большой кривизне дуги аорты. [195]
Источник
Последовательность пре-мРНК miR-712 генерируется из мышиного рибосомального гена RN45s во внутреннем транскрибируемом спейсерном регионе 2 (ITS2). [195] XRN1 — это экзонуклеаза, которая разрушает регион ITS2 во время обработки RN45s. [195] Таким образом, уменьшение XRN1 в условиях d-flow приводит к накоплению miR-712. [195]
Механизм
MiR-712 нацелен на тканевой ингибитор металлопротеиназ 3 (TIMP3). [195] TIMP обычно регулируют активность матриксных металлопротеиназ (MMP), которые разрушают внеклеточный матрикс (ECM). Артериальный ECM в основном состоит из коллагеновых и эластиновых волокон, обеспечивающих структурную поддержку и свойства отдачи артерий. [196] Эти волокна играют важную роль в регуляции сосудистого воспаления и проницаемости, которые важны для развития атеросклероза. [197] Экспрессируемый эндотелиальными клетками, TIMP3 является единственным TIMP, связанным с ECM. [196] Снижение экспрессии TIMP3 приводит к увеличению деградации ECM в присутствии d-потока. В соответствии с этими результатами, ингибирование pre-miR712 увеличивает экспрессию TIMP3 в клетках, даже при воздействии турбулентного потока. [195]
TIMP3 также снижает экспрессию TNFα (провоспалительного регулятора) во время турбулентного потока. [195] Активность TNFα в турбулентном потоке измерялась по экспрессии TNFα-превращающего фермента (TACE) в крови. TNFα снижался, если miR-712 был ингибирован или TIMP3 был сверхэкспрессирован, [195] предполагая, что miR-712 и TIMP3 регулируют активность TACE в условиях турбулентного потока.
Анти-miR-712 эффективно подавляет экспрессию miR-712, вызванную d-flow, и увеличивает экспрессию TIMP3. [195] Анти-miR-712 также подавляет сосудистую гиперпроницаемость, тем самым значительно снижая развитие атеросклеротических поражений и инфильтрацию иммунных клеток. [195]
Человеческий гомолог микроРНК-205
Человеческий гомолог miR-712 был обнаружен в гене-гомологе RN45s, который поддерживает сходные miRNA с мышами. [195] MiR-205 человека имеет схожие последовательности с miR-712 мышей и сохраняется у большинства позвоночных. [195] MiR-205 и miR-712 также разделяют более 50% мишеней клеточной сигнализации, включая TIMP3. [195]
При тестировании d-flow снизил экспрессию XRN1 у людей, как и в эндотелиальных клетках мышей, что указывает на потенциально распространенную роль XRN1 у людей. [195]
Болезнь почек
Целенаправленное удаление Dicer в почечных клетках-предшественниках, полученных из FoxD1, в мышиной модели привело к сложному почечному фенотипу, включающему расширение предшественников нефронов , меньшее количество клеток ренина , артериолы гладких мышц , прогрессирующую потерю мезангии и аневризмы клубочков. [198] Высокопроизводительное профилирование всего транскриптома мышиной модели с нокаутом FoxD1-Dicer выявило эктопическую регуляцию проапоптотического гена Bcl2L11 (Bim) и нарушение регуляции пути p53 с увеличением эффекторных генов p53, включая Bax , Trp53inp1 , Jun, Cdkn1a , Mmp2 и Arid3a . Уровни белка p53 остались неизменными, что позволяет предположить, что стромальные miRNA FoxD1 напрямую подавляют эффекторные гены p53. Используя подход отслеживания линий с последующей сортировкой клеток с использованием флуоресцентной активации , профилирование miRNA клеток, полученных от FoxD1, не только всесторонне определило транскрипционный ландшафт miRNA, имеющих решающее значение для развития сосудов, но и идентифицировало ключевые miRNA, которые, вероятно, будут модулировать почечный фенотип в его отсутствие. Эти miRNA включают miRs-10a, 18a, 19b, 24, 30c, 92a, 106a, 130a, 152, 181a, 214, 222, 302a, 370 и 381, которые регулируют Bcl2L11 (Bim), и miRs-15b, 18a, 21, 30c, 92a, 106a, 125b-5p, 145, 214, 222, 296-5p и 302a, которые регулируют гены-эффекторы p53. В соответствии с результатами профилирования, эктопический апоптоз наблюдался в клеточных производных линии предшественников, полученной от FoxD1, и подтверждает важность почечных стромальных микроРНК в клеточном гомеостазе. [198]
Нервная система
МикроРНК имеют решающее значение для здорового развития и функционирования нервной системы . [199] Предыдущие исследования показывают, что микроРНК могут регулировать нейрональную дифференцировку и созревание на различных стадиях. [200] МикроРНК также играют важную роль в синаптическом развитии [201] (например, в дендритогенезе или морфогенезе шипиков) и синаптической пластичности [202] (способствуя обучению и памяти). Устранение образования микроРНК у мышей путем экспериментального подавления Dicer привело к патологическим результатам, таким как уменьшение размера нейронов, двигательные аномалии (при подавлении в нейронах полосатого тела [203] ) и нейродегенерация (при подавлении в нейронах переднего мозга [204] ). Измененная экспрессия miRNA была обнаружена при нейродегенеративных заболеваниях (таких как болезнь Альцгеймера , болезнь Паркинсона и болезнь Хантингтона [205] ), а также при многих психиатрических расстройствах (включая эпилепсию , [206] шизофрению , большую депрессию , биполярное расстройство и тревожные расстройства [207] [208] [209] ).
Гладить
По данным Центра по контролю и профилактике заболеваний, инсульт является одной из основных причин смерти и длительной инвалидности в Америке. 87% случаев — это ишемические инсульты , которые возникают из-за закупорки артерии мозга, которая несет богатую кислородом кровь. Обструкция кровотока означает, что мозг не может получать необходимые питательные вещества, такие как кислород и глюкоза, и удалять отходы, такие как углекислый газ. [210] [211] miRNAs играет роль в посттрансляционном подавлении генов, нацеливаясь на гены в патогенезе церебральной ишемии, такие как воспалительный, ангиогенез и апоптотический путь. [212]
Алкоголизм
Жизненно важная роль miRNA в экспрессии генов имеет важное значение для наркомании , в частности алкоголизма . [213] Хроническое злоупотребление алкоголем приводит к стойким изменениям в работе мозга, частично опосредованным изменениями в экспрессии генов . [213] Глобальная регуляция miRNA многих нижестоящих генов считается значимой в отношении реорганизации синаптических связей или долгосрочных нейронных адаптаций, включающих изменение поведения от потребления алкоголя до отмены и/или зависимости . [214] Было обнаружено, что до 35 различных miRNA были изменены в мозге алкоголика после смерти, все из которых нацелены на гены, которые включают регуляцию клеточного цикла , апоптоз , клеточную адгезию , развитие нервной системы и клеточную сигнализацию . [213] Измененные уровни miRNA были обнаружены в медиальной префронтальной коре мышей, зависимых от алкоголя, что предполагает роль miRNA в организации трансляционного дисбаланса и создании дифференциально экспрессируемых белков в области мозга, где, вероятно, возникают сложное когнитивное поведение и принятие решений . [215]
miRNAs могут быть как повышены, так и понижены в ответ на хроническое употребление алкоголя. Экспрессия miR-206 увеличилась в префронтальной коре у крыс, зависимых от алкоголя, нацеливаясь на фактор транскрипции нейротрофического фактора мозга ( BDNF ) и в конечном итоге снижая его экспрессию. BDNF играет важную роль в формировании и созревании новых нейронов и синапсов, что предполагает возможное участие в росте синапсов/ синаптической пластичности у лиц, злоупотребляющих алкоголем. [216] Было обнаружено, что miR-155 , важный в регуляции реакций нейровоспаления , вызванных алкоголем , повышен, что предполагает роль микроглии и воспалительных цитокинов в патофизиологии алкоголя. [217] Понижение экспрессии miR-382 было обнаружено в прилежащем ядре , структуре в базальном переднем мозге, важной для регуляции чувств вознаграждения , которые питают мотивационные привычки. miR-382 является мишенью для дофаминового рецептора D1 (DRD1), и его сверхэкспрессия приводит к повышению регуляции DRD1 и delta fosB , фактора транскрипции, который активирует серию событий транскрипции в прилежащем ядре , что в конечном итоге приводит к аддиктивному поведению. [218] Альтернативно, сверхэкспрессия miR-382 привела к ослаблению употребления алкоголя и ингибированию повышения регуляции DRD1 и delta fosB в моделях алкоголизма на крысах, что демонстрирует возможность использования фармацевтических препаратов, нацеленных на miRNA, в лечении. [218]
Ожирение
miRNAs играют решающую роль в регуляции предшественников стволовых клеток , дифференцирующихся в адипоциты . [219] Исследования, направленные на определение роли плюрипотентных стволовых клеток в адипогенезе , были изучены на линии стромальных клеток, полученных из костного мозга человека hMSC-Tert20. [220] Снижение экспрессии miR-155 , miR-221 и miR-222 было обнаружено во время адипогенного программирования как иммортализованных, так и первичных hMSC, что позволяет предположить, что они действуют как отрицательные регуляторы дифференциации. Напротив, эктопическая экспрессия miRNAs 155 , 221 и 222 значительно ингибировала адипогенез и подавляла индукцию главных регуляторов PPARγ и CCAAT/энхансер-связывающего белка альфа ( CEBPA ). [221] Это открывает путь для возможных генетических методов лечения ожирения.
Другой класс miRNA, регулирующих резистентность к инсулину , ожирение и диабет , — это семейство let-7 . Let-7 накапливается в тканях человека в процессе старения . [222] Когда let-7 был эктопически сверхэкспрессирован для имитации ускоренного старения, мыши стали резистентными к инсулину и, таким образом, более склонными к ожирению и диабету , вызванному диетой с высоким содержанием жиров . [223] Напротив, когда let-7 был ингибирован инъекциями антагомиров , специфичных для let-7 , мыши стали более чувствительными к инсулину и значительно устойчивыми к ожирению и диабету, вызванному диетой с высоким содержанием жиров. Ингибирование let-7 может не только предотвратить ожирение и диабет, но и обратить вспять и вылечить это состояние. [224] Эти экспериментальные результаты свидетельствуют о том, что ингибирование let-7 может представлять собой новую терапию ожирения и диабета 2 типа.
Гемостаз
miRNA также играют важную роль в регуляции сложных ферментативных каскадов, включая гемостатическую систему свертывания крови . [225] Крупномасштабные исследования функционального нацеливания miRNA недавно выявили рациональные терапевтические цели в гемостатической системе. [226] [227] Они были напрямую связаны с гомеостазом кальция в эндоплазматическом ретикулуме , который имеет решающее значение для дифференциации клеток на ранних стадиях развития. [228]
Растения
miRNA считаются ключевыми регуляторами многих процессов развития, гомеостаза и иммунных процессов в растениях. [229] Их роль в развитии растений включает развитие апикальной меристемы побега , рост листьев, образование цветков, производство семян или расширение корней. [230] [231] [232] [233] Кроме того, они играют сложную роль в ответах на различные абиотические стрессы , включая тепловой стресс, низкотемпературный стресс, стресс от засухи, световой стресс или воздействие гамма-излучения. [229]
miRNA могут связываться с целевыми транскриптами РНК-мессенджера (мРНК) генов, кодирующих белки, и негативно контролировать их трансляцию или вызывать деградацию мРНК. Крайне важно точно идентифицировать цели miRNA. [237] Доступно сравнение предсказательной эффективности восемнадцати алгоритмов in silico . [238] Масштабные исследования функционального нацеливания miRNA показывают, что многие функциональные miRNA могут быть пропущены алгоритмами прогнозирования целей. [226]
^ abc Bartel DP (январь 2004 г.). «МикроРНК: геномика, биогенез, механизм и функция». Cell . 116 (2): 281–297. doi : 10.1016/S0092-8674(04)00045-5 . PMID 14744438.
^ Куреши А, Такур Н, Монга И, Такур А, Кумар М (январь 2014 г.). "VIRmiRNA: всеобъемлющий ресурс для экспериментально подтвержденных вирусных miRNA и их целей". База данных . 2014 : bau103. doi :10.1093/database/bau103. PMC 4224276. PMID 25380780 .
^ abc Bartel DP (январь 2009 г.). «МикроРНК: распознавание целей и регуляторные функции». Cell . 136 (2): 215–233. doi :10.1016/j.cell.2009.01.002. PMC 3794896 . PMID 19167326.
^ Jonas S, Izaurralde E (июль 2015 г.). «На пути к молекулярному пониманию подавления генов, опосредованного микроРНК». Nature Reviews. Genetics . 16 (7): 421–433. doi :10.1038/nrg3965. PMID 26077373. S2CID 24892348.
^ Jonas S, Izaurralde E (июль 2015 г.). «На пути к молекулярному пониманию подавления генов, опосредованного микроРНК». Nature Reviews. Genetics . 16 (7): 421–433. doi :10.1038/nrg3965. PMID 26077373. S2CID 24892348.
^ Alles J, Fehlmann T, Fischer U, Backes C, Galata V, Minet M и др. (апрель 2019 г.). «Оценка общего числа истинных человеческих miRNA». Nucleic Acids Research . 47 (7): 3353–3364. doi :10.1093/nar/gkz097. PMC 6468295. PMID 30820533 .{{cite journal}}: CS1 maint: переопределенная настройка ( ссылка )
^ Фромм Б., Доманска Д., Хойе Э., Овчинников В., Канг В., Апарисио-Пуэрта Э. и др. (январь 2020 г.). «MirGeneDB 2.0: Комплемент микроРНК многоклеточных животных». Исследования нуклеиновых кислот . 48 (Д1): Д132–Д141. дои : 10.1093/nar/gkz885. ПМК 6943042 . ПМИД 31598695.{{cite journal}}: CS1 maint: переопределенная настройка ( ссылка )
^ Лим Л.П., Лау Н.К., Вайнштейн Э.Г., Абдельхаким А., Йекта С., Роудс М.В. и др. (апрель 2003 г.). «МикроРНК Caenorhabditis elegans». Гены и развитие . 17 (8): 991–1008. дои : 10.1101/gad.1074403. ПМК 196042 . ПМИД 12672692.{{cite journal}}: CS1 maint: переопределенная настройка ( ссылка )
^ ab Lagos-Quintana M, Rauhut R, Yalcin A, Meyer J, Lendeckel W, Tuschl T (апрель 2002 г.). «Идентификация тканеспецифичных микроРНК у мышей». Current Biology . 12 (9): 735–739. Bibcode :2002CBio...12..735L. doi : 10.1016/S0960-9822(02)00809-6 . PMID 12007417.
^ abc Lewis BP, Burge CB, Bartel DP (январь 2005 г.). «Консервативное спаривание семян, часто фланкированное аденозинами, указывает на то, что тысячи человеческих генов являются мишенями микроРНК». Cell . 120 (1): 15–20. doi : 10.1016/j.cell.2004.12.035 . PMID 15652477.
^ abc Friedman RC, Farh KK, Burge CB, Bartel DP (январь 2009 г.). «Большинство мРНК млекопитающих являются консервативными мишенями микроРНК». Genome Research . 19 (1): 92–105. doi :10.1101/gr.082701.108. PMC 2612969 . PMID 18955434.
^ Fromm B, Billipp T, Peck LE, Johansen M, Tarver JE, King BL и др. (2015). «Единая система для аннотации генов микроРНК позвоночных и эволюция микроРНКома человека». Annual Review of Genetics . 49 (1): 213–242. Bibcode : 2015ARGen..49..213F. doi : 10.1146/annurev-genet-120213-092023. PMC 4743252. PMID 26473382 .{{cite journal}}: CS1 maint: переопределенная настройка ( ссылка )
^ «Нобелевская премия по физиологии и медицине 2024 года».
^ "Нобелевская премия по физиологии и медицине 2024 года". NobelPrize.org (пресс-релиз) . Получено 7 октября 2024 г.
^ Льюис Т. (7 октября 2024 г.). «Нобелевская премия по физиологии и медицине присуждена за открытие регуляции генов микроРНК». Scientific American . Получено 7 октября 2024 г.
^ abc Lee RC, Feinbaum RL, Ambros V (декабрь 1993 г.). "Гетерохронный ген C. elegans lin-4 кодирует малые РНК с антисмысловой комплементарностью к lin-14". Cell . 75 (5): 843–54. doi : 10.1016/0092-8674(93)90529-Y . PMID 8252621.
^ ab Reinhart BJ, Slack FJ, Basson M, Pasquinelli AE, Bettinger JC, Rougvie AE и др. (февраль 2000 г.). «21-нуклеотидная let-7 РНК регулирует сроки развития у Caenorhabditis elegans». Nature . 403 (6772): 901–6. Bibcode :2000Natur.403..901R. doi :10.1038/35002607. PMID 10706289. S2CID 4384503.
^ ab Pasquinelli AE, Reinhart BJ, Slack F, Martindale MQ, Kuroda MI, Maller B, et al. (Ноябрь 2000). "Консервация последовательности и временной экспрессии гетерохронной регуляторной РНК let-7". Nature . 408 (6808): 86–9. Bibcode :2000Natur.408...86P. doi :10.1038/35040556. PMID 11081512. S2CID 4401732.
^ abc Lagos-Quintana M, Rauhut R, Lendeckel W, Tuschl T (октябрь 2001 г.). «Идентификация новых генов, кодирующих малые экспрессируемые РНК». Science . 294 (5543): 853–8. Bibcode :2001Sci...294..853L. doi :10.1126/science.1064921. hdl : 11858/00-001M-0000-0012-F65F-2 . PMID 11679670. S2CID 18101169.
^ abc Lau NC, Lim LP, Weinstein EG, Bartel DP (октябрь 2001 г.). «Обильный класс крошечных РНК с вероятными регуляторными ролями у Caenorhabditis elegans». Science . 294 (5543): 858–62. Bibcode :2001Sci...294..858L. doi :10.1126/science.1065062. PMID 11679671. S2CID 43262684.
^ abc Lee RC, Ambros V (октябрь 2001 г.). «Обширный класс малых РНК у Caenorhabditis elegans». Science . 294 (5543): 862–4. Bibcode :2001Sci...294..862L. doi :10.1126/science.1065329. PMID 11679672. S2CID 33480585.
^ Винхольдс Э., Клоостерман В.П., Миска Э., Альварес-Сааведра Э., Березиков Э., де Брёйн Э. и др. (июль 2005 г.). «Экспрессия микроРНК в эмбриональном развитии рыбок данио». Наука . 309 (5732): 310–1. Бибкод : 2005Sci...309..310W. дои : 10.1126/science.1114519. PMID 15919954. S2CID 38939571.
^ ab Jones-Rhoades MW, Bartel DP, Bartel B (2006). «МикроРНК и их регуляторные роли в растениях». Annual Review of Plant Biology . 57 : 19–53. doi :10.1146/annurev.arplant.57.032905.105218. PMID 16669754.
^ Brennecke J, Hipfner DR, Stark A, Russell RB, Cohen SM (апрель 2003 г.). «bantam кодирует регулируемую развитием микроРНК, которая контролирует пролиферацию клеток и регулирует проапоптотический ген hid у Drosophila». Cell . 113 (1): 25–36. doi : 10.1016/S0092-8674(03)00231-9 . PMID 12679032.
^ Cuellar TL, McManus MT (декабрь 2005 г.). «МикроРНК и эндокринная биология». Журнал эндокринологии . 187 (3): 327–32. doi : 10.1677/joe.1.06426 . PMID 16423811.
^ Poy MN, Eliasson L, Krutzfeldt J, Kuwajima S, Ma X, Macdonald PE и др. (ноябрь 2004 г.). «МикроРНК, специфичная для панкреатических островков, регулирует секрецию инсулина». Nature . 432 (7014): 226–30. Bibcode :2004Natur.432..226P. doi :10.1038/nature03076. PMID 15538371. S2CID 4415988.
^ Wilfred BR, Wang WX, Nelson PT (июль 2007 г.). «Энергетическое исследование miRNA: обзор роли miRNA в метаболизме липидов с предсказанием, что miR-103/107 регулирует метаболические пути человека». Молекулярная генетика и метаболизм . 91 (3): 209–17. doi :10.1016/j.ymgme.2007.03.011. PMC 1978064. PMID 17521938 .
^ Harfe BD, McManus MT, Mansfield JH, Hornstein E, Tabin CJ (август 2005 г.). «Фермент РНКазы III Dicer необходим для морфогенеза, но не для формирования паттерна конечности позвоночного». Труды Национальной академии наук Соединенных Штатов Америки . 102 (31): 10898–903. Bibcode : 2005PNAS..10210898H. doi : 10.1073/pnas.0504834102 . PMC 1182454. PMID 16040801 .
^ Trang P, Weidhaas JB, Slack FJ (декабрь 2008 г.). «МикроРНК как потенциальные средства лечения рака». Oncogene . 27 (Suppl 2): S52–7. doi : 10.1038/onc.2009.353 . PMC 10033140 . PMID 19956180.
^ Li C, Feng Y, Coukos G, Zhang L (декабрь 2009 г.). «Стратегии терапевтической микроРНК при раке человека». Журнал AAPS . 11 (4): 747–57. doi :10.1208/s12248-009-9145-9. PMC 2782079. PMID 19876744 .
^ Fasanaro P, Greco S, Ivan M, Capogrossi MC, Martelli F (январь 2010 г.). «микроРНК: новые терапевтические мишени при острых ишемических заболеваниях». Pharmacology & Therapeutics . 125 (1): 92–104. doi :10.1016/j.pharmthera.2009.10.003. PMID 19896977.
^ Hydbring P, Badalian-Very G (август 2013 г.). "Клиническое применение микроРНК". F1000Research . 2 : 136. doi : 10.12688/f1000research.2-136.v2 . PMC 3917658. PMID 24627783 .
^ Wightman B, Ha I, Ruvkun G (декабрь 1993 г.). "Посттранскрипционная регуляция гетерохронного гена lin-14 с помощью lin-4 опосредует формирование временного паттерна у C. elegans". Cell . 75 (5): 855–62. doi : 10.1016/0092-8674(93)90530-4 . PMID 8252622.
^ Giza DE, Calin GA (2015). «МикроРНК и хронический лимфоцитарный лейкоз». МикроРНК: Рак . Достижения в экспериментальной медицине и биологии. Том 889. С. 23–40. doi :10.1007/978-3-319-23730-5_2. ISBN978-3-319-23729-9. PMID 26658994.
^ Ambros V, Bartel B, Bartel DP, Burge CB, Carrington JC, Chen X и др. (март 2003 г.). «Единая система аннотации микроРНК». RNA . 9 (3): 277–9. doi :10.1261/rna.2183803. PMC 1370393 . PMID 12592000.
^ Griffiths-Jones S, Grocock RJ, van Dongen S, Bateman A, Enright AJ (январь 2006 г.). "miRBase: последовательности микроРНК, цели и номенклатура генов". Nucleic Acids Research . 34 (выпуск базы данных): D140–4. doi :10.1093/nar/gkj112. PMC 1347474. PMID 16381832 .
^ Wright MW, Bruford EA (январь 2011 г.). «Название „мусора“: номенклатура генов человеческой некодирующей РНК (нкРНК)». Human Genomics . 5 (2): 90–8. doi : 10.1186/1479-7364-5-2-90 . PMC 3051107 . PMID 21296742.
^ Хант М., Банерджи С., Сурана П., Лю М., Фюрст Г., Матиони С. и др. (2019). «Открытие малых РНК во взаимодействии ячменя и возбудителя мучнистой росы». BMC Genomics . 20 (1): 19–53. doi : 10.1186/s12864-019-5947-z . PMC 6657096. PMID 31345162 .
^ abc Lewis BP, Shih IH, Jones-Rhoades MW, Bartel DP, Burge CB (декабрь 2003 г.). «Прогнозирование целей микроРНК млекопитающих». Cell . 115 (7): 787–98. doi : 10.1016/S0092-8674(03)01018-3 . PMID 14697198.
^ Ellwanger DC, Büttner FA, Mewes HW, Stümpflen V (май 2011 г.). «Достаточный минимальный набор типов семян miRNA». Биоинформатика . 27 (10): 1346–50. doi :10.1093/bioinformatics/btr149. PMC 3087955. PMID 21441577 .
^ Rajewsky N (июнь 2006 г.). "микроРНК-целевые предсказания у животных". Nature Genetics . 38 (6s): S8–13. doi :10.1038/ng1798. PMID 16736023. S2CID 23496396.
^ Крек А., Грюн Д., Пой М.Н., Вольф Р., Розенберг Л., Эпштейн Э.Дж. и др. (май 2005 г.). «Комбинаторные предсказания целевых микроРНК». Nature Genetics . 37 (5): 495–500. doi :10.1038/ng1536. PMID 15806104. S2CID 22672750.
^ Thomson DW, Bracken CP, Goodall GJ (сентябрь 2011 г.). «Экспериментальные стратегии идентификации мишеней микроРНК». Nucleic Acids Research . 39 (16): 6845–53. doi : 10.1093/nar/gkr330. PMC 3167600. PMID 21652644.
^ ab Lim LP, Lau NC, Garrett-Engele P, Grimson A, Schelter JM, Castle J, et al. (февраль 2005 г.). «Анализ микрочипов показывает, что некоторые микроРНК подавляют большое количество целевых мРНК». Nature . 433 (7027): 769–73. Bibcode :2005Natur.433..769L. doi :10.1038/nature03315. PMID 15685193. S2CID 4430576.
^ Selbach M, Schwanhäusser B, Thierfelder N, Fang Z, Khanin R, Rajewsky N (сентябрь 2008 г.). «Широко распространенные изменения в синтезе белка, вызванные микроРНК». Nature . 455 (7209): 58–63. Bibcode :2008Natur.455...58S. doi :10.1038/nature07228. PMID 18668040. S2CID 4429008.
^ Baek D, Villén J, Shin C, Camargo FD, Gygi SP, Bartel DP (сентябрь 2008 г.). «Влияние микроРНК на выход белка». Nature . 455 (7209): 64–71. Bibcode :2008Natur.455...64B. doi :10.1038/nature07242. PMC 2745094 . PMID 18668037.
^ ab Rodriguez A, Griffiths-Jones S, Ashurst JL, Bradley A (октябрь 2004 г.). «Идентификация генов-хозяев и единиц транскрипции микроРНК млекопитающих». Genome Research . 14 (10A): 1902–10. doi :10.1101/gr.2722704. PMC 524413 . PMID 15364901.
^ abcd Cai X, Hagedorn CH, Cullen BR (декабрь 2004 г.). «Человеческие микроРНК обрабатываются из кэпированных полиаденилированных транскриптов, которые также могут функционировать как мРНК». РНК . 10 (12): 1957–66. doi :10.1261/rna.7135204. PMC 1370684 . PMID 15525708.
^ Weber MJ (январь 2005 г.). «Новые гены микроРНК человека и мыши, найденные путем поиска гомологии». Журнал FEBS . 272 (1): 59–73. doi : 10.1111/j.1432-1033.2004.04389.x . PMID 15634332. S2CID 32923462.
^ Ким YK, Ким VN (февраль 2007). «Обработка интронных микроРНК». The EMBO Journal . 26 (3): 775–83. doi :10.1038/sj.emboj.7601512. PMC 1794378. PMID 17255951 .
^ Baskerville S, Bartel DP (март 2005 г.). «Профилирование микроРНК с помощью микрочипов выявляет частую коэкспрессию с соседними микроРНК и генами хозяина». РНК . 11 (3): 241–7. doi :10.1261/rna.7240905. PMC 1370713 . PMID 15701730.
^ abc Lee Y, Kim M, Han J, Yeom KH, Lee S, Baek SH и др. (октябрь 2004 г.). «Гены микроРНК транскрибируются РНК-полимеразой II». The EMBO Journal . 23 (20): 4051–60. doi :10.1038/sj.emboj.7600385. PMC 524334. PMID 15372072 .
^ Zhou X, Ruan J, Wang G, Zhang W (март 2007 г.). "Характеристика и идентификация основных промоторов микроРНК в четырех модельных видах". PLOS Computational Biology . 3 (3): e37. Bibcode : 2007PLSCB...3...37Z. doi : 10.1371/journal.pcbi.0030037 . PMC 1817659. PMID 17352530 .
^ Faller M, Guo F (ноябрь 2008 г.). «Биогенез микроРНК: есть больше, чем один способ снять шкуру с кошки». Biochimica et Biophysica Acta (BBA) - Механизмы регуляции генов . 1779 (11): 663–7. doi :10.1016/j.bbagrm.2008.08.005. PMC 2633599. PMID 18778799 .
^ Ли Ю, Ан С, Хан Дж, Чой Х, Ким Дж, Йим Дж и др. (сентябрь 2003 г.). «Ядерная РНКаза III Дроша инициирует процессинг микроРНК». Природа . 425 (6956): 415–9. Бибкод : 2003Natur.425..415L. дои : 10.1038/nature01957. PMID 14508493. S2CID 4421030.
^ Gregory RI, Chendrimada TP, Shiekhattar R (2006). «Биогенез микроРНК: изоляция и характеристика микропроцессорного комплекса». Протоколы микроРНК . Методы в молекулярной биологии. Т. 342. С. 33–47. doi :10.1385/1-59745-123-1:33. ISBN978-1-59745-123-9. PMID 16957365.
^ Han J, Lee Y, Yeom KH, Kim YK, Jin H, Kim VN (декабрь 2004 г.). «Комплекс Drosha-DGCR8 в первичной обработке микроРНК». Genes & Development . 18 (24): 3016–27. doi :10.1101/gad.1262504. PMC 535913. PMID 15574589 .
^ Han J, Lee Y, Yeom KH, Nam JW, Heo I, Rhee JK и др. (июнь 2006 г.). «Молекулярная основа распознавания первичных микроРНК комплексом Drosha-DGCR8». Cell . 125 (5): 887–901. doi : 10.1016/j.cell.2006.03.043 . PMID 16751099.
^ Conrad T, Marsico A, Gehre M, Orom UA (октябрь 2014 г.). «Активность микропроцессора контролирует дифференциальный биогенез miRNA in Vivo». Cell Reports . 9 (2): 542–54. doi : 10.1016/j.celrep.2014.09.007 . PMID 25310978.
^ Auyeung VC, Ulitsky I, McGeary SE, Bartel DP (февраль 2013 г.). «За пределами вторичной структуры: детерминанты первичной последовательности лицензируют шпильки pri-miRNA для процессинга». Cell . 152 (4): 844–58. doi :10.1016/j.cell.2013.01.031. PMC 3707628 . PMID 23415231.
^ Ali PS, Ghoshdastider U, Hoffmann J, Brutschy B, Filipek S (ноябрь 2012 г.). «Распознавание предшественника miRNA let-7g человеческим Lin28B». FEBS Letters . 586 (22): 3986–90. Bibcode : 2012FEBSL.586.3986S. doi : 10.1016/j.febslet.2012.09.034 . PMID 23063642. S2CID 28899778.
^ ab Ruby JG, Jan CH, Bartel DP (июль 2007 г.). «Предшественники интронных микроРНК, обходящие обработку Drosha». Nature . 448 (7149): 83–86. Bibcode :2007Natur.448...83R. doi :10.1038/nature05983. PMC 2475599 . PMID 17589500.
^ Березиков Э., Чунг В.Дж., Уиллис Дж., Куппен Э., Лай ЕС (октябрь 2007 г.). «Миртронные гены млекопитающих». Молекулярная клетка . 28 (2): 328–336. doi :10.1016/j.molcel.2007.09.028. ПМЦ 2763384 . ПМИД 17964270.
^ аб Кавахара Ю., Меграу М., Крайдер Э., Иисаса Х., Валенте Л., Хацигеоргиу А.Г. и др. (сентябрь 2008 г.). «Частота и судьба редактирования микроРНК в мозге человека». Исследования нуклеиновых кислот . 36 (16): 5270–80. дои : 10.1093/nar/gkn479. ПМК 2532740 . ПМИД 18684997.
^ Winter J, Jung S, Keller S, Gregory RI, Diederichs S (март 2009). «Множество дорог к зрелости: пути биогенеза микроРНК и их регуляция». Nature Cell Biology . 11 (3): 228–34. doi :10.1038/ncb0309-228. PMID 19255566. S2CID 205286318.
^ Ohman M (октябрь 2007 г.). «Претендент на редактирование A-to-I или союзник процесса микроРНК». Biochimie . 89 (10): 1171–6. doi :10.1016/j.biochi.2007.06.002. PMID 17628290.
^ ab Murchison EP , Hannon GJ (июнь 2004 г.). "miRNAs on the move: miRNA biogenesis and the RNAi machinery". Current Opinion in Cell Biology . 16 (3): 223–9. doi :10.1016/j.ceb.2004.04.003. PMID 15145345.
^ abc Lund E, Dahlberg JE (2006). «Субстратная селективность экспортина 5 и Dicer в биогенезе микроРНК». Симпозиумы Cold Spring Harbor по количественной биологии . 71 : 59–66. doi : 10.1101/sqb.2006.71.050 . PMID 17381281.
^ Park JE, Heo I, Tian Y, Simanshu DK, Chang H, Jee D и др. (июль 2011 г.). «Dicer распознает 5'-конец РНК для эффективной и точной обработки». Nature . 475 (7355): 201–5. doi :10.1038/nature10198. PMC 4693635 . PMID 21753850.
^ Ji X (2008). «Механизм действия РНКазы III: как Dicer Dices». РНК-интерференция . Текущие темы микробиологии и иммунологии. Том 320. С. 99–116. doi :10.1007/978-3-540-75157-1_5. ISBN978-3-540-75156-4. PMID 18268841.
^ Mirihana Arachchilage G, Dassanayake AC, Basu S (февраль 2015 г.). «Структурный переключатель РНК, зависимый от ионов калия, регулирует созревание пре-микроРНК 92b человека». Химия и биология . 22 (2): 262–72. doi : 10.1016/j.chembiol.2014.12.013 . PMID 25641166.
^ Sohel MH (2016). «Внеклеточные/циркулирующие микроРНК: механизмы высвобождения, функции и проблемы». Достижения в области наук о жизни . 10 (2): 175–186. doi : 10.1016/j.als.2016.11.007 .
^ ab Boeckel JN, Reis SM, Leistner D, Thomé CE, Zeiher AM, Fichtlscherer S, et al. (апрель 2014 г.). «От сердца до кончиков пальцев ног: вклад сердца в периферические уровни микроРНК». Международный журнал кардиологии . 172 (3): 616–7. doi :10.1016/j.ijcard.2014.01.082. PMID 24508494.
^ Lelandais-Brière C, Sorin C, Declerck M, Benslimane A, Crespi M, Hartmann C (март 2010 г.). «Разнообразие малых РНК в растениях и его влияние на развитие». Current Genomics . 11 (1): 14–23. doi :10.2174/138920210790217918. PMC 2851111 . PMID 20808519.
^ Rana TM (январь 2007 г.). «Освещая тишину: понимание структуры и функции малых РНК». Nature Reviews Molecular Cell Biology . 8 (1): 23–36. doi :10.1038/nrm2085. PMID 17183358. S2CID 8966239.
^ ab Schwarz DS, Zamore PD (май 2002). «Почему miRNAs живут в miRNP?». Genes & Development . 16 (9): 1025–31. doi : 10.1101/gad.992502 . PMID 12000786.
^ Krol J, Sobczak K, Wilczynska U, Drath M, Jasinska A, Kaczynska D и др. (октябрь 2004 г.). «Структурные особенности предшественников микроРНК (миРНК) и их значение для биогенеза микроРНК и дизайна малых интерферирующих РНК/коротких шпилечных РНК». Журнал биологической химии . 279 (40): 42230–9. doi : 10.1074/jbc.M404931200 . PMID 15292246.
^ Хворова А., Рейнольдс А., Джаясена С.Д. (октябрь 2003 г.). «Функциональные siRNA и miRNA проявляют смещение цепи». Cell . 115 (2): 209–16. doi : 10.1016/S0092-8674(03)00801-8 . PMID 14567918.
^ Schwarz DS, Hutvágner G, Du T, Xu Z, Aronin N, Zamore PD (октябрь 2003 г.). «Асимметрия в сборке комплекса ферментов РНК-интерференции». Cell . 115 (2): 199–208. doi : 10.1016/S0092-8674(03)00759-1 . PMID 14567917.
^ Lin SL, Chang D, Ying SY (август 2005 г.). «Асимметрия интронных пре-микроРНК-структур в функциональной сборке RISC». Gene . 356 : 32–8. doi :10.1016/j.gene.2005.04.036. PMC 1788082 . PMID 16005165.
^ Okamura K, Chung WJ, Lai EC (сентябрь 2008 г.). «Длинные и короткие инвертированные повторные гены у животных: микроРНК, миртроны и шпильковые РНК». Cell Cycle . 7 (18): 2840–5. doi :10.4161/cc.7.18.6734. PMC 2697033 . PMID 18769156.
^ ab Pratt AJ, MacRae IJ (июль 2009 г.). «Комплекс подавления экспрессии, индуцированный РНК: универсальная машина подавления экспрессии генов». Журнал биологической химии . 284 (27): 17897–901. doi : 10.1074/jbc.R900012200 . PMC 2709356. PMID 19342379 .
^ MacRae IJ, Ma E, Zhou M, Robinson CV, Doudna JA (январь 2008 г.). «In vitro reconstitution of the human RISC-loading complex». Труды Национальной академии наук Соединенных Штатов Америки . 105 (2): 512–7. Bibcode : 2008PNAS..105..512M. doi : 10.1073/pnas.0710869105 . PMC 2206567. PMID 18178619 .
^ Mourelatos Z, Dostie J, Paushkin S, Sharma A, Charroux B, Abel L, et al. (март 2002 г.). «miRNP: новый класс рибонуклеопротеинов, содержащих многочисленные микроРНК». Genes & Development . 16 (6): 720–8. doi :10.1101/gad.974702. PMC 155365 . PMID 11914277.
^ Meister G, Landthaler M, Peters L, Chen PY, Urlaub H, Lührmann R и др. (декабрь 2005 г.). «Идентификация новых белков, связанных с аргонавтами». Current Biology . 15 (23): 2149–55. Bibcode :2005CBio...15.2149M. doi : 10.1016/j.cub.2005.10.048 . hdl : 11858/00-001M-0000-0012-E763-B . PMID 16289642.
^ Shaw G, Kamen R (29 августа 1986 г.). «Консервативная последовательность AU из 3'-нетранслируемой области мРНК GM-CSF опосредует селективную деградацию мРНК». Cell . 46 (5): 659–667. doi :10.1016/0092-8674(86)90341-7. ISSN 0092-8674. PMID 3488815.
^ Jing Q, Huang S, Guth S, Zarubin T, Motoyama A, Chen J, et al. (март 2005 г.). «Участие микроРНК в нестабильности мРНК, опосредованной богатыми AU элементами». Cell . 120 (5): 623–34. doi : 10.1016/j.cell.2004.12.038 . PMID 15766526.
^ Wang XJ, Reyes JL, Chua NH, Gaasterland T (2004). «Прогнозирование и идентификация микроРНК Arabidopsis thaliana и их мишеней мРНК». Genome Biology . 5 (9): R65. doi : 10.1186/gb-2004-5-9-r65 . PMC 522872. PMID 15345049 .
^ Кавасаки Х, Тайра К (2004). «МикроРНК-196 ингибирует экспрессию HOXB8 при миелоидной дифференцировке клеток HL60». Серия симпозиумов по нуклеиновым кислотам . 48 (1): 211–2. doi : 10.1093/nass/48.1.211 . PMID 17150553.
^ ab Moxon S, Jing R, Szittya G, Schwach F, Rusholme Pilcher RL, Moulton V и др. (октябрь 2008 г.). «Глубокое секвенирование коротких РНК томатов выявляет микроРНК, нацеленные на гены, участвующие в созревании плодов». Genome Research . 18 (10): 1602–9. doi :10.1101/gr.080127.108. PMC 2556272 . PMID 18653800.
^ Mazière P, Enright AJ (июнь 2007 г.). «Предсказание целей микроРНК». Drug Discovery Today . 12 (11–12): 452–8. doi :10.1016/j.drudis.2007.04.002. PMID 17532529.
^ Williams AE (февраль 2008 г.). «Функциональные аспекты животных микроРНК». Cellular and Molecular Life Sciences . 65 (4): 545–62. doi :10.1007/s00018-007-7355-9. PMC 11131689. PMID 17965831. S2CID 5708394 .
^ Эулалио А., Ханцингер Э., Нишихара Т., Ревинкель Дж., Фаузер М., Изаурральде Э. (январь 2009 г.). «Деаденилирование - широко распространенный эффект регуляции микроРНК». РНК . 15 (1): 21–32. дои : 10.1261/rna.1399509. ПМЦ 2612776 . ПМИД 19029310.
^ Bazzini AA, Lee MT, Giraldez AJ (апрель 2012 г.). «Профилирование рибосом показывает, что miR-430 снижает трансляцию, прежде чем вызвать распад мРНК у данио-рерио». Science . 336 (6078): 233–7. Bibcode :2012Sci...336..233B. doi :10.1126/science.1215704. PMC 3547538 . PMID 22422859.
^ Djuranovic S, Nahvi A, Green R (апрель 2012 г.). «опосредованное miRNA подавление генов путем трансляционной репрессии с последующим деаденилированием и распадом мРНК». Science . 336 (6078): 237–40. Bibcode :2012Sci...336..237D. doi :10.1126/science.1215691. PMC 3971879 . PMID 22499947.
^ Tan Y, Zhang B, Wu T, Skogerbø G, Zhu X, Guo X и др. (февраль 2009 г.). «Транскрипционное ингибирование экспрессии Hoxd4 miRNA-10a в клетках рака молочной железы человека». BMC Molecular Biology . 10 (1): 12. doi : 10.1186/1471-2199-10-12 . PMC 2680403 . PMID 19232136.
^ Stark A, Brennecke J, Bushati N, Russell RB, Cohen SM (декабрь 2005 г.). «Животные микроРНК придают устойчивость экспрессии генов и оказывают значительное влияние на эволюцию 3'UTR». Cell . 123 (6): 1133–46. doi : 10.1016/j.cell.2005.11.023 . PMID 16337999.
^ He L, Hannon GJ (июль 2004 г.). «МикроРНК: малые РНК с большой ролью в регуляции генов». Nature Reviews. Genetics . 5 (7): 522–531. doi :10.1038/nrg1379. PMID 15211354. S2CID 5270062.
^ Li LC (2008). «Активация генов, опосредованная малыми РНК». В Morris KV (ред.). РНК и регуляция экспрессии генов: скрытый уровень сложности. Horizon Scientific Press. ISBN978-1-904455-25-7.
^ Place RF, Li LC, Pookot D, Noonan EJ, Dahiya R (февраль 2008 г.). «МикроРНК-373 индуцирует экспрессию генов с комплементарными промоторными последовательностями». Труды Национальной академии наук Соединенных Штатов Америки . 105 (5): 1608–13. Bibcode : 2008PNAS..105.1608P. doi : 10.1073/pnas.0707594105 . PMC 2234192. PMID 18227514 .(Опечатка: doi :10.1073/pnas.1803343115, PMID 29555737, Retraction Watch . Если опечатка была проверена и не влияет на цитируемый материал, замените на . ){{erratum|...}}{{erratum|...|checked=yes}}
^ ab Kumar S, Reddy PH (сентябрь 2016 г.). «Являются ли циркулирующие микроРНК периферическими биомаркерами болезни Альцгеймера?». Biochim Biophys Acta . 1862 (9): 1617–27. doi :10.1016/j.bbadis.2016.06.001. PMC 5343750. PMID 27264337 .
^ van den Berg MM, Krauskopf J, Ramaekers JG и др. (февраль 2020 г.). «Циркулирующие микроРНК как потенциальные биомаркеры психиатрических и нейродегенеративных расстройств». Prog Neurobiol . 185 : 101732. doi : 10.1016/j.pneurobio.2019.101732 . PMID 31816349.
^ Cuman C, Van Sinderen M, Gantier MP, Rainczuk K, Sorby K, Rombauts L и др. (октябрь 2015 г.). «Секретируемая бластоцистой микроРНК регулирует адгезию эндометриальных эпителиальных клеток». eBioMedicine . 2 (10): 1528–1535. doi :10.1016/j.ebiom.2015.09.003. PMC 4634783 . PMID 26629549.
^ Zhou L, Miller C, Miraglia LJ, Romero A, Mure LS, Panda S и др. (январь 2021 г.). «Полногеномный скрининг микроРНК идентифицирует кластер микроРНК-183/96/182 как модулятор циркадных ритмов». Труды Национальной академии наук Соединенных Штатов Америки . 118 (1): e2020454118. Bibcode : 2021PNAS..11820454Z. doi : 10.1073/pnas.2020454118 . PMC 7817116. PMID 33443164. S2CID 230713808 .
«МикроРНК играют ключевую роль в регуляции циркадных ритмов». Новости науки . 6 января 2021 г.
^ Axtell MJ, Bartel DP (июнь 2005 г.). «Древность микроРНК и их мишени в наземных растениях». The Plant Cell . 17 (6): 1658–73. doi : 10.1105 /tpc.105.032185. PMC 1143068. PMID 15849273.
^ Tanzer A, Stadler PF (май 2004). «Молекулярная эволюция кластера микроРНК». Журнал молекулярной биологии . 339 (2): 327–35. CiteSeerX 10.1.1.194.1598 . doi :10.1016/j.jmb.2004.03.065. PMID 15136036.
^ Чен К, Раевский Н (февраль 2007 г.). «Эволюция регуляции генов факторами транскрипции и микроРНК». Nature Reviews Genetics . 8 (2): 93–103. doi :10.1038/nrg1990. PMID 17230196. S2CID 174231.
^ Lee CT, Risom T, Strauss WM (апрель 2007 г.). «Эволюционная консервация регуляторных цепей микроРНК: исследование сложности генов микроРНК и консервативных взаимодействий микроРНК-мишень через филогению метазоа». ДНК и клеточная биология . 26 (4): 209–18. doi :10.1089/dna.2006.0545. PMID 17465887.
^ abcd Peterson KJ, Dietrich MR, McPeek MA (июль 2009 г.). «МикроРНК и макроэволюция метазойных организмов: взгляд на канализацию, сложность и кембрийский взрыв». BioEssays . 31 (7): 736–47. doi : 10.1002/bies.200900033 . PMID 19472371. S2CID 15364875.
^ Шабалина СА, Кунин ЕВ (октябрь 2008). "Истоки и эволюция эукариотической РНК-интерференции". Trends in Ecology & Evolution . 23 (10): 578–87. Bibcode :2008TEcoE..23..578S. doi :10.1016/j.tree.2008.06.005. PMC 2695246. PMID 18715673 .
^ Axtell MJ, Westholm JO, Lai EC (2011). "Vive la différence: биогенез и эволюция микроРНК у растений и животных". Genome Biology . 12 (4): 221. doi : 10.1186/gb-2011-12-4-221 . PMC 3218855. PMID 21554756 .
^ ab Wheeler BM, Heimberg AM, Moy VN, Sperling EA, Holstein TW, Heber S, et al. (2009). «Глубокая эволюция микроРНК метазойных организмов». Эволюция и развитие . 11 (1): 50–68. doi :10.1111/j.1525-142X.2008.00302.x. PMID 19196333. S2CID 14924603.
^ Пашковский ПП, Рязанский СС (июнь 2013). «Биогенез, эволюция и функции микроРНК растений». Биохимия. Биохимия . 78 (6): 627–37. doi :10.1134/S0006297913060084. PMID 23980889. S2CID 12025420.
^ ab Heimberg AM, Sempere LF, Moy VN, Donoghue PC, Peterson KJ (февраль 2008 г.). «МикроРНК и возникновение морфологической сложности позвоночных». Труды Национальной академии наук Соединенных Штатов Америки . 105 (8): 2946–50. Bibcode : 2008PNAS..105.2946H. doi : 10.1073/pnas.0712259105 . PMC 2268565. PMID 18287013 .
^ abc Nozawa M, Miura S, Nei M (июль 2010 г.). «Происхождение и эволюция генов микроРНК у видов Drosophila». Genome Biology and Evolution . 2 : 180–9. doi :10.1093/gbe/evq009. PMC 2942034 . PMID 20624724.
^ Allen E, Xie Z, Gustafson AM, Sung GH, Spatafora JW, Carrington JC (декабрь 2004 г.). «Эволюция генов микроРНК путем инвертированной дупликации последовательностей целевых генов в Arabidopsis thaliana». Nature Genetics . 36 (12): 1282–90. doi :10.1038/ng1478. PMID 15565108. S2CID 11997028.
^ Warthmann N, Das S, Lanz C, Weigel D (май 2008). «Сравнительный анализ локуса микроРНК MIR319a у Arabidopsis и родственных Brassicaceae». Молекулярная биология и эволюция . 25 (5): 892–902. doi : 10.1093/molbev/msn029 . PMID 18296705.
^ Фальгрен Н., Джогдео С., Касшау К.Д., Салливан С.М., Чепмен Э.Дж., Лаубингер С. и др. (апрель 2010 г.). «Эволюция генов микроРНК у Arabidopsis lyrata и Arabidopsis thaliana». Растительная клетка . 22 (4): 1074–89. дои : 10.1105/tpc.110.073999. ПМЦ 2879733 . ПМИД 20407027.
^ Caravas J, Friedrich M (июнь 2010 г.). «О клещах и многоножках: недавний прогресс в разрешении основы дерева членистоногих». BioEssays . 32 (6): 488–95. doi :10.1002/bies.201000005. PMID 20486135. S2CID 20548122.
^ Kenny NJ, Namigai EK, Marlétaz F, Hui JH, Shimeld SM (декабрь 2015 г.). «Черновик геномных сборок и предсказанные микроРНК-комплементы литоральных лофотрохозоев Patella vulgata (Mollusca, Patellogastropoda) и Spirobranchus (Pomatoceros) lamarcki (Annelida, Serpulida)». Marine Genomics . 24 (2): 139–46. Bibcode :2015MarGn..24..139K. doi :10.1016/j.margen.2015.07.004. PMID 26319627.
^ Cock JM, Sterck L, Rouzé P, Scornet D, Allen AE, Amoutzias G, et al. (Июнь 2010 г.). «Геном Ectocarpus и независимая эволюция многоклеточности у бурых водорослей». Nature . 465 (7298): 617–21. Bibcode :2010Natur.465..617C. doi : 10.1038/nature09016 . PMID 20520714.
^ Cuperus JT, Fahlgren N, Carrington JC (февраль 2011 г.). «Эволюция и функциональная диверсификация генов MIRNA». The Plant Cell . 23 (2): 431–42. doi :10.1105/tpc.110.082784. PMC 3077775. PMID 21317375 .
^ Райан Дж. Ф., Панг К., Шницлер CE, Нгуен АД, Морленд РТ, Симмонс Д. К. и др. (декабрь 2013 г.). «Геном гребневика Mnemiopsis leidyi и его значение для эволюции типа клеток». Science . 342 (6164): 1242592. doi :10.1126/science.1242592. PMC 3920664 . PMID 24337300.
^ Maxwell EK, Ryan JF, Schnitzler CE, Browne WE, Baxevanis AD (декабрь 2012 г.). «МикроРНК и основные компоненты механизма обработки микроРНК не кодируются в геноме гребневика Mnemiopsis leidyi». BMC Genomics . 13 (1): 714. doi : 10.1186/1471-2164-13-714 . PMC 3563456 . PMID 23256903.
^ Dimond PF (15 марта 2010 г.). "Терапевтический потенциал miRNAs". Новости генной инженерии и биотехнологии . 30 (6): 1. Архивировано из оригинала 19 июля 2010 г. Получено 10 июля 2010 г.
^ Tjaden B, Goodwin SS, Opdyke JA, Guillier M, Fu DX, Gottesman S и др. (2006). «Предсказание цели для малых некодирующих РНК в бактериях». Nucleic Acids Research . 34 (9): 2791–802. doi :10.1093/nar/gkl356. PMC 1464411. PMID 16717284 .
^ Liu CG, Calin GA, Volinia S, Croce CM (2008). «Профилирование экспрессии микроРНК с использованием микрочипов». Nature Protocols . 3 (4): 563–78. doi :10.1038/nprot.2008.14. PMID 18388938. S2CID 2441105.
^ Chen C, Ridzon DA, Broomer AJ, Zhou Z, Lee DH, Nguyen JT и др. (ноябрь 2005 г.). «Количественная оценка микроРНК в реальном времени методом ОТ-ПЦР с использованием стебель-петли». Nucleic Acids Research . 33 (20): e179. doi :10.1093/nar/gni178. PMC 1292995. PMID 16314309 .
^ Shingara J, Keiger K, Shelton J, Laosinchai-Wolf W, Powers P, Conrad R и др. (сентябрь 2005 г.). «Оптимизированная платформа изоляции и маркировки для точного профилирования экспрессии микроРНК». RNA . 11 (9): 1461–70. doi :10.1261/rna.2610405. PMC 1370829 . PMID 16043497.
^ Buermans HP, Ariyurek Y, van Ommen G, den Dunnen JT, 't Hoen PA (декабрь 2010 г.). «Новые методы для профилирования экспрессии микроРНК на основе секвенирования следующего поколения». BMC Genomics . 11 : 716. doi : 10.1186/1471-2164-11-716 . PMC 3022920. PMID 21171994 .
^ Kloosterman WP, Wienholds E, Ketting RF, Plasterk RH (2004). «Требования к субстрату для функции let-7 в развивающемся эмбрионе данио-рерио». Nucleic Acids Research . 32 (21): 6284–91. doi :10.1093/nar/gkh968. PMC 535676. PMID 15585662 .
^ Flynt AS, Li N, Thatcher EJ, Solnica-Krezel L, Patton JG (февраль 2007 г.). «Zebrafish miR-214 модулирует сигнализацию Hedgehog, чтобы определить судьбу мышечных клеток». Nature Genetics . 39 (2): 259–63. doi :10.1038/ng1953. PMC 3982799 . PMID 17220889.
^ Meister G, Landthaler M, Dorsett Y, Tuschl T (март 2004 г.). «Специфическое ингибирование подавления РНК, вызванного микроРНК и siRNA». РНК . 10 (3): 544–50. doi :10.1261/rna.5235104. PMC 1370948 . PMID 14970398.
^ Kloosterman WP, Lagendijk AK, Ketting RF, Moulton JD, Plasterk RH (август 2007 г.). «Целенаправленное ингибирование созревания miRNA с помощью морфолино выявляет роль miR-375 в развитии островков поджелудочной железы». PLOS Biology . 5 (8): e203. doi : 10.1371/journal.pbio.0050203 . PMC 1925136 . PMID 17676975.(В настоящее время в этой статье выражена обеспокоенность , см. doi :10.1371/journal.pbio.3001631, PMID 35486652. Если это преднамеренная ссылка на такую статью, замените на . ){{expression of concern|...}}{{expression of concern|...|intentional=yes}}
^ Gebert LF, Rebhan MA, Crivelli SE, Denzler R, Stoffel M, Hall J (январь 2014 г.). «Miravirsen (SPC3649) может ингибировать биогенез miR-122». Nucleic Acids Research . 42 (1): 609–21. doi :10.1093/nar/gkt852. PMC 3874169. PMID 24068553 .
^ Choi WY, Giraldez AJ, Schier AF (октябрь 2007 г.). «Защитники мишеней выявляют подавление и балансировку узловых агонистов и антагонистов с помощью miR-430». Science . 318 (5848): 271–4. Bibcode :2007Sci...318..271C. doi : 10.1126/science.1147535 . PMID 17761850. S2CID 30461594.
^ You Y, Moreira BG, Behlke MA, Owczarzy R (май 2006 г.). «Разработка зондов LNA, улучшающих дискриминацию несоответствий». Nucleic Acids Research . 34 (8): e60. doi :10.1093/nar/gkl175. PMC 1456327. PMID 16670427 .
^ Lagendijk AK, Moulton JD, Bakkers J (июнь 2012 г.). «Раскрытие деталей: протокол гибридизации in situ целых образцов микроРНК для эмбрионов данио-рерио и тканей взрослых животных». Biology Open . 1 (6): 566–9. doi :10.1242/bio.2012810. PMC 3509442. PMID 23213449 .
^ Kaur H, Arora A, Wengel J, Maiti S (июнь 2006 г.). «Термодинамические, противоионные и гидратационные эффекты для включения запертых нуклеотидов нуклеиновых кислот в дуплексы ДНК». Биохимия . 45 (23): 7347–55. doi :10.1021/bi060307w. PMID 16752924.
^ Nielsen JA, Lau P, Maric D, Barker JL, Hudson LD (август 2009 г.). «Интеграция профилей экспрессии микроРНК и мРНК нейрональных предшественников для выявления регуляторных сетей, лежащих в основе начала кортикального нейрогенеза». BMC Neuroscience . 10 : 98. doi : 10.1186/1471-2202-10-98 . PMC 2736963 . PMID 19689821.
^ Grimson A, Farh KK, Johnston WK, Garrett-Engele P, Lim LP, Bartel DP (июль 2007 г.). «Специфичность нацеливания микроРНК у млекопитающих: детерминанты, выходящие за рамки спаривания семян». Molecular Cell . 27 (1): 91–105. doi :10.1016/j.molcel.2007.06.017. PMC 3800283 . PMID 17612493.
^ Гриффитс-Джонс С., Сайни Х.К., ван Донген С., Энрайт А.Дж. (январь 2008 г.). "miRBase: инструменты для геномики микроРНК". Nucleic Acids Research . 36 (выпуск базы данных): D154–8. doi :10.1093/nar/gkm952. PMC 2238936. PMID 17991681 .
^ Nam S, Li M, Choi K, Balch C, Kim S, Nephew KP (июль 2009 г.). «Интегрированный анализ микроРНК и мРНК (MMIA): веб-инструмент для изучения биологических функций экспрессии микроРНК». Nucleic Acids Research . 37 (выпуск веб-сервера): W356–62. doi :10.1093/nar/gkp294. PMC 2703907. PMID 19420067 .
^ Artmann S, Jung K, Bleckmann A, Beissbarth T (2012). Provero P (ред.). «Обнаружение одновременных групповых эффектов в экспрессии микроРНК и связанных целевых наборах генов». PLOS ONE . 7 (6): e38365. Bibcode :2012PLoSO...738365A. doi : 10.1371/journal.pone.0038365 . PMC 3378551 . PMID 22723856.
^ Jiang Q, Wang Y, Hao Y, Juan L, Teng M, Zhang X и др. (январь 2009 г.). "miR2Disease: вручную курируемая база данных по дерегуляции микроРНК при заболеваниях человека". Nucleic Acids Research . 37. 37 (выпуск базы данных): D98–104. doi :10.1093/nar/gkn714. PMC 2686559 . PMID 18927107.
^ Mencía A, Modamio-Høybjør S, Redshaw N, Morín M, Mayo-Merino F, Olavarrieta L и др. (май 2009 г.). «Мутации в области семян человеческой miR-96 ответственны за несиндромную прогрессирующую потерю слуха». Nature Genetics . 41 (5): 609–13. doi :10.1038/ng.355. PMID 19363479. S2CID 11113852.
^ Hughes AE, Bradley DT, Campbell M, Lechner J, Dash DP, Simpson DA и др. (ноябрь 2011 г.). «Мутация, изменяющая область семян miR-184, вызывает семейный кератоконус с катарактой». American Journal of Human Genetics . 89 (5): 628–33. doi :10.1016/j.ajhg.2011.09.014. PMC 3213395 . PMID 21996275.
^ де Понтуаль Л., Яо Э., Калье П., Фавр Л., Друэн В., Кариу С. и др. (сентябрь 2011 г.). «Удаление зародышевой линии кластера миР-17~92 вызывает дефекты скелета и роста у людей». Природная генетика . 43 (10): 1026–30. дои : 10.1038/ng.915. ПМК 3184212 . ПМИД 21892160.
^ Calin GA, Liu CG, Sevignani C, Ferracin M, Felli N, Dumitru CD и др. (август 2004 г.). «Профилирование микроРНК выявляет отличительные признаки хронических лимфоцитарных лейкемий В-клеток». Труды Национальной академии наук Соединенных Штатов Америки . 101 (32): 11755–11760. Bibcode : 2004PNAS..10111755C. doi : 10.1073/pnas.0404432101 . PMC 511048. PMID 15284443 .
^ Velle A, Pesenti C, Grassi T, Beltrame L, Martini P, Jaconi M и др. (Май 2023 г.). «Комплексное исследование гистотип-специфических микроРНК и их вариантов при эпителиальном раке яичников I стадии». International Journal of Cancer . 152 (9): 1989–2001. doi : 10.1002/ijc.34408 . hdl : 11577/3478062 . PMID 36541726. S2CID 255034585.
^ Кириакидис И, Кириакидис К, Цезу А (август 2022 г.). «МикроРНК и диагностика острого лимфобластного лейкоза у детей: систематический обзор, метаанализ и повторный анализ с использованием новых инструментов секвенирования малых РНК». Раковые заболевания . 14 (16): 3976. doi : 10.3390/cancers14163976 . PMC 9406077. PMID 36010971 .
^ Võsa U, Vooder T, Kolde R, Fischer K, Välk K, Tõnisson N и др. (октябрь 2011 г.). «Идентификация miR-374a как прогностического маркера выживаемости у пациентов с ранней стадией немелкоклеточного рака легких». Genes, Chromosomes & Cancer . 50 (10): 812–22. doi :10.1002/gcc.20902. PMID 21748820. S2CID 9746594.
^ Akçakaya P, Ekelund S, Kolosenko I, Caramuta S, Ozata DM, Xie H и др. (август 2011 г.). «Нарушение регуляции miR-185 и miR-133b связано с общей выживаемостью и метастазами при колоректальном раке». International Journal of Oncology . 39 (2): 311–8. doi : 10.3892/ijo.2011.1043 . PMID 21573504.
^ ab Eyking A, Reis H, Frank M, Gerken G, Schmid KW, Cario E (2016). «MiR-205 и MiR-373 связаны с агрессивным муцинозным колоректальным раком человека». PLOS ONE . 11 (6): e0156871. Bibcode : 2016PLoSO..1156871E. doi : 10.1371/journal.pone.0156871 . PMC 4894642. PMID 27271572 .
^ МикроРНК-21 способствует пролиферации клеток гепатоцеллюлярной карциномы HepG2 посредством подавления митоген-активируемой протеинкиназы-киназы 3. Гуансянь Сюй и др., 2013
^ Jones K, Nourse JP, Keane C, Bhatnagar A, Gandhi MK (январь 2014 г.). «Плазменные микроРНК являются биомаркерами ответа на заболевание при классической лимфоме Ходжкина». Clinical Cancer Research . 20 (1): 253–64. doi : 10.1158/1078-0432.CCR-13-1024 . PMID 24222179.
^ Hosseinahli N, Aghapour M, Duijf PH, Baradaran B (август 2018 г.). «Лечение рака с помощью заместительной терапии микроРНК: обзор литературы». Журнал клеточной физиологии . 233 (8): 5574–5588. doi : 10.1002/jcp.26514 . PMID 29521426. S2CID 3766576.
^ Liu G, Sun Y, Ji P, Li X, Cogdell D, Yang D и др. (июль 2014 г.). «MiR-506 подавляет пролиферацию и индуцирует старение, напрямую воздействуя на ось CDK4/6-FOXM1 при раке яичников». Журнал патологии . 233 (3): 308–18. doi :10.1002/path.4348. PMC 4144705. PMID 24604117 .
^ Wen SY, Lin Y, Yu YQ, Cao SJ, Zhang R, Yang XM и др. (февраль 2015 г.). «miR-506 действует как супрессор опухолей, напрямую воздействуя на фактор транскрипции пути hedgehog Gli3 при раке шейки матки у человека». Oncogene . 34 (6): 717–25. doi : 10.1038/onc.2014.9 . PMID 24608427. S2CID 20603801.
↑ Пекинский университет (13 сентября 2022 г.). «Ученые разрабатывают новое мощное оружие для борьбы с раком». Cell . 185 (11). SciTech Daily : 1888–1904.e24. doi : 10.1016/j.cell.2022.04.030 . PMID 35623329. S2CID 249070106. Получено 15 сентября 2022 г.
^ Loeb KR, Loeb LA (март 2000). «Значение множественных мутаций при раке». Канцерогенез . 21 (3): 379–385. doi : 10.1093/carcin/21.3.379 . PMID 10688858.
^ Lodish H, Berk A, Kaiser CA, Krieger M, Bretscher A, Ploegh H, et al. (2016). Молекулярная клеточная биология (8-е изд.). Нью-Йорк: WH Freeman and Company. стр. 203. ISBN978-1-4641-8339-3.
^ Hu H, Gatti RA (июнь 2011 г.). «МикроРНК: новые игроки в реакции на повреждение ДНК». Журнал молекулярной клеточной биологии . 3 (3): 151–158. doi : 10.1093/jmcb/mjq042. PMC 3104011. PMID 21183529.
^ Jasperson KW, Tuohy TM, Neklason DW, Burt RW (июнь 2010 г.). «Наследственный и семейный рак толстой кишки». Гастроэнтерология . 138 (6): 2044–58. doi :10.1053/j.gastro.2010.01.054. PMC 3057468. PMID 20420945 .
^ Truninger K, Menigatti M, Luz J, Russell A, Haider R, Gebbers JO и др. (май 2005 г.). «Иммуногистохимический анализ выявляет высокую частоту дефектов PMS2 при колоректальном раке». Гастроэнтерология . 128 (5): 1160–71. doi : 10.1053/j.gastro.2005.01.056 . PMID 15887099.
^ Валери Н., Гаспарини П., Фаббри М., Бракони К., Веронезе А., Ловат Ф. и др. (апрель 2010 г.). «Модуляция репарации несоответствий и геномной стабильности с помощью miR-155». Труды Национальной академии наук Соединенных Штатов Америки . 107 (15): 6982–7. Bibcode : 2010PNAS..107.6982V. doi : 10.1073/pnas.1002472107 . PMC 2872463. PMID 20351277 .
^ abc Zhang W, Zhang J, Hoadley K, Kushwaha D, Ramakrishnan V, Li S, et al. (июнь 2012 г.). "miR-181d: предиктивный биомаркер глиобластомы, который подавляет экспрессию MGMT". Neuro-Oncology . 14 (6): 712–9. doi :10.1093/neuonc/nos089. PMC 3367855 . PMID 22570426.
^ Spiegl-Kreinecker S, Pirker C, Filipits M, Lötsch D, Buchroithner J, Pichler J, et al. (Январь 2010 г.). «Экспрессия белка O6-метилгуанин ДНК метилтрансферазы в опухолевых клетках предсказывает исход терапии темозоломидом у пациентов с глиобластомой». Neuro-Oncology . 12 (1): 28–36. doi :10.1093/neuonc/nop003. PMC 2940563 . PMID 20150365.
^ Сгарра Р., Рустиги А., Тессари М.А., Ди Бернардо Дж., Альтамура С., Фуско А. и др. (сентябрь 2004 г.). «Ядерные фосфопротеины HMGA и их связь со структурой хроматина и раком». Письма ФЭБС . 574 (1–3): 1–8. Бибкод : 2004FEBSL.574....1S. doi :10.1016/j.febslet.2004.08.013. PMID 15358530. S2CID 28903539.
^ Xu Y, Sumter TF, Bhattacharya R, Tesfaye A, Fuchs EJ, Wood LJ и др. (май 2004 г.). «Онкоген HMG-I вызывает высокопенетрантную, агрессивную лимфоидную злокачественность у трансгенных мышей и сверхэкспрессируется при лейкемии человека». Cancer Research . 64 (10): 3371–5. doi : 10.1158/0008-5472.CAN-04-0044 . PMID 15150086.
^ Borrmann L, Schwanbeck R, Heyduk T, Seebeck B, Rogalla P, Bullerdiek J, et al. (декабрь 2003 г.). «Высокомобильный белок группы A2 и его производные связывают специфическую область промотора гена репарации ДНК ERCC1 и модулируют его активность». Nucleic Acids Research . 31 (23): 6841–51. doi :10.1093/nar/gkg884. PMC 290254 . PMID 14627817.
^ Facista A, Nguyen H, Lewis C, Prasad AR, Ramsey L, Zaitlin B и др. (апрель 2012 г.). «Недостаточная экспрессия ферментов репарации ДНК при раннем прогрессировании спорадического рака толстой кишки». Genome Integrity . 3 (1): 3. doi : 10.1186/2041-9414-3-3 . PMC 3351028. PMID 22494821.
^ Gibert B, Delloye-Bourgeois C, Gattolliat CH, Meurette O, Le Guernevel S, Fombonne J, et al. (Ноябрь 2014 г.). «Регулирование семейством miR181 подавления опухолей рецептора CDON при нейробластоме». Журнал Национального института рака . 106 (11). doi : 10.1093/jnci/dju318 . PMID 25313246.
^ Chen JF, Murchison EP , Tang R, Callis TE, Tatsuguchi M, Deng Z и др. (февраль 2008 г.). «Целенаправленное удаление Dicer в сердце приводит к дилатационной кардиомиопатии и сердечной недостаточности». Труды Национальной академии наук Соединенных Штатов Америки . 105 (6): 2111–6. Bibcode : 2008PNAS..105.2111C. doi : 10.1073/pnas.0710228105 . PMC 2542870. PMID 18256189 .
^ ab Zhao Y, Ransom JF, Li A, Vedantham V, von Drehle M, Muth AN и др. (апрель 2007 г.). «Нарушение регуляции кардиогенеза, сердечной проводимости и клеточного цикла у мышей с отсутствием miRNA-1-2». Cell . 129 (2): 303–17. doi : 10.1016/j.cell.2007.03.030 . PMID 17397913.
^ Thum T, Galuppo P, Wolf C, Fiedler J, Kneitz S, van Laake LW и др. (Июль 2007 г.). «МикроРНК в человеческом сердце: ключ к перепрограммированию генов плода при сердечной недостаточности». Circulation . 116 (3): 258–67. doi : 10.1161/CIRCULATIONAHA.107.687947 . PMID 17606841.
^ van Rooij E, Sutherland LB, Liu N, Williams AH, McAnally J, Gerard RD и др. (ноябрь 2006 г.). «Сигнатурный паттерн стресс-чувствительных микроРНК, которые могут вызывать гипертрофию сердца и сердечную недостаточность». Труды Национальной академии наук Соединенных Штатов Америки . 103 (48): 18255–60. Bibcode : 2006PNAS..10318255V. doi : 10.1073 /pnas.0608791103 . PMC 1838739. PMID 17108080.
^ Tatsuguchi M, Seok HY, Callis TE, Thomson JM, Chen JF, Newman M и др. (июнь 2007 г.). «Экспрессия микроРНК динамически регулируется во время гипертрофии кардиомиоцитов». Журнал молекулярной и клеточной кардиологии . 42 (6): 1137–41. doi :10.1016/j.yjmcc.2007.04.004. PMC 1934409. PMID 17498736 .
^ Zhao Y, Samal E, Srivastava D (июль 2005 г.). «Фактор ответа сыворотки регулирует специфичную для мышц микроРНК, которая воздействует на Hand2 во время кардиогенеза». Nature . 436 (7048): 214–20. Bibcode :2005Natur.436..214Z. doi :10.1038/nature03817. PMID 15951802. S2CID 4340449.
^ Xiao J, Luo X, Lin H, Zhang Y, Lu Y, Wang N и др. (апрель 2007 г.). «МикроРНК miR-133 подавляет экспрессию канала HERG K+, способствуя удлинению QT в диабетических сердцах». Журнал биологической химии . 282 (17): 12363–7. doi : 10.1074/jbc.C700015200 . PMC 3151106. PMID 17344217.(Отозвано, см. doi :10.1074/jbc.A111.700015, PMID 21961173, Retraction Watch . Если это преднамеренная ссылка на отозванную статью, замените на . ){{retracted|...}}{{retracted|...|intentional=yes}}
^ Yang B, Lin H, Xiao J, Lu Y, Luo X, Li B и др. (апрель 2007 г.). «Мышечно-специфическая микроРНК miR-1 регулирует сердечный аритмогенный потенциал, воздействуя на GJA1 и KCNJ2». Nature Medicine . 13 (4): 486–91. doi :10.1038/nm1569. PMID 17401374. S2CID 1935811.
^ Каре А., Каталуччи Д., Феличетти Ф., Бончи Д., Аддарио А., Галло П. и др. (май 2007 г.). «МикроРНК-133 контролирует гипертрофию сердца». Природная медицина . 13 (5): 613–8. дои : 10.1038/nm1582. PMID 17468766. S2CID 10097893.
^ van Rooij E, Sutherland LB, Qi X, Richardson JA, Hill J, Olson EN (апрель 2007 г.). «Контроль стресс-зависимого сердечного роста и экспрессии генов с помощью микроРНК». Science . 316 (5824): 575–9. Bibcode :2007Sci...316..575V. doi :10.1126/science.1139089. PMID 17379774. S2CID 1927839.
^ Keller T, Boeckel JN, Groß S, Klotsche J, Palapies L, Leistner D и др. (Июль 2017 г.). «Улучшенная стратификация риска в профилактике с использованием панели выбранных циркулирующих микроРНК». Scientific Reports . 7 (1): 4511. Bibcode :2017NatSR...7.4511K. doi :10.1038/s41598-017-04040-w. PMC 5495799 . PMID 28674420.
^ Insull W (январь 2009). «Патология атеросклероза: развитие бляшек и реакция бляшек на медицинское лечение». Американский журнал медицины . 122 (1 Suppl): S3–S14. doi :10.1016/j.amjmed.2008.10.013. PMID 19110086.
^ abcdefghijklmnopq Son DJ, Kumar S, Takabe W, Kim CW, Ni CW, Alberts-Grill N и др. (2013). «Атипичная механочувствительная микроРНК-712, полученная из прерибосомальной РНК, вызывает воспаление эндотелия и атеросклероз». Nature Communications . 4 : 3000. Bibcode :2013NatCo...4.3000S. doi :10.1038/ncomms4000. PMC 3923891 . PMID 24346612.
^ ab Basu R, Fan D, Kandalam V, Lee J, Das SK, Wang X и др. (декабрь 2012 г.). «Потеря гена Timp3 приводит к образованию аневризмы брюшной аорты в ответ на ангиотензин II». Журнал биологической химии . 287 (53): 44083–96. doi : 10.1074/jbc.M112.425652 . PMC 3531724. PMID 23144462 .
^ Либби П. (2002). «Воспаление при атеросклерозе». Nature . 420 (6917): 868–74. Bibcode : 2002Natur.420..868L. doi : 10.1038/nature01323. PMID 12490960. S2CID 407449.
^ ab Phua YL, Chu JY, Marrone AK, Bodnar AJ, Sims-Lucas S, Ho J (октябрь 2015 г.). «Почечные стромальные miRNAs необходимы для нормального нефрогенеза и выживания мезангиальных клубочков». Physiological Reports . 3 (10): e12537. doi :10.14814/phy2.12537. PMC 4632944 . PMID 26438731.
^ Cao DD, Li L, Chan WY (май 2016 г.). «МикроРНК: ключевые регуляторы в центральной нервной системе и их роль в неврологических заболеваниях». Международный журнал молекулярных наук . 17 (6): 842. doi : 10.3390/ijms17060842 . PMC 4926376. PMID 27240359 .
^ Lang MF, Shi Y (2012). «Динамические роли микроРНК в нейрогенезе». Frontiers in Neuroscience . 6 : 71. doi : 10.3389/fnins.2012.00071 . PMC 3356852. PMID 22661924 .
^ Schratt G (декабрь 2009 г.). «микроРНК в синапсе». Nature Reviews. Neuroscience . 10 (12): 842–849. doi :10.1038/nrn2763. PMID 19888283. S2CID 3507952.
^ Luo M, Li L, Ding M, Niu Y, Xu X, Shi X и др. (19 января 2023 г.). «Длительное потенцирование и регуляторные микроРНК депрессии были выделены при ухудшении обучения и памяти, вызванном бисфенолом А, с помощью секвенирования микроРНК и биоинформатического анализа». PLOS ONE . 18 (1): e0279029. Bibcode :2023PLoSO..1879029L. doi : 10.1371/journal.pone.0279029 . PMC 9851566 . PMID 36656826.
^ Cuellar TL, Davis TH, Nelson PT, Loeb GB, Harfe BD, Ullian E и др. (апрель 2008 г.). «Потеря Dicer в нейронах полосатого тела приводит к поведенческим и нейроанатомическим фенотипам при отсутствии нейродегенерации». Труды Национальной академии наук Соединенных Штатов Америки . 105 (14): 5614–5619. Bibcode : 2008PNAS..105.5614C. doi : 10.1073/pnas.0801689105 . PMC 2291142. PMID 18385371 .
^ Эбер СС, Пападопулу А.С., Смит П., Галас М.К., Планель Э., Силахтароглу А.Н. и др. (октябрь 2010 г.). «Генетическая абляция Dicer в нейронах переднего мозга взрослых приводит к аномальному гиперфосфорилированию тау и нейродегенерации». Молекулярная генетика человека . 19 (20): 3959–3969. дои : 10.1093/hmg/ddq311 . ПМИД 20660113.
^ Рой Б., Ли Э., Ли Т., Рамперсо М. (февраль 2022 г.). «Роль микроРНК в нейродегенерации: от причины заболевания до инструментов обнаружения биомаркеров и терапии». Гены . 13 ( 3): 425. doi : 10.3390/genes13030425 . PMC 8951370. PMID 35327979.
^ Henshall DC, Hamer HM, Pasterkamp RJ, Goldstein DB, Kjems J, Prehn JH и др. (декабрь 2016 г.). «МикроРНК при эпилепсии: патофизиология и клиническая полезность». The Lancet. Неврология . 15 (13): 1368–1376. doi : 10.1016/S1474-4422(16)30246-0 . PMID 27839653.
^ Hommers LG, Domschke K, Deckert J (январь 2015 г.). «Гетерогенность и индивидуальность: микроРНК при психических расстройствах» (PDF) . Journal of Neural Transmission . 122 (1): 79–97. doi :10.1007/s00702-014-1338-4. PMID 25395183. S2CID 25088900 . Архивировано (PDF) из оригинала 23 мая 2022 г.
^ Feng J, Sun G, Yan J, Noltner K, Li W, Buzin CH и др. (июль 2009 г.). Reif A (ред.). «Доказательства шизофрении X-хромосомы, связанной с изменениями микроРНК». PLOS ONE . 4 (7): e6121. Bibcode : 2009PLoSO...4.6121F . doi : 10.1371/journal.pone.0006121 . PMC 2699475 . PMID 19568434.
^ Beveridge NJ, Gardiner E, Carroll AP, Tooney PA, Cairns MJ (декабрь 2010 г.). «Шизофрения связана с увеличением биогенеза кортикальной микроРНК». Молекулярная психиатрия . 15 (12): 1176–1189. doi : 10.1038/mp.2009.84 . PMC 2990188. PMID 19721432.
^ "Факты об инсульте". Центры по контролю и профилактике заболеваний . 15 марта 2019 г. Получено 5 декабря 2019 г.
^ Rink C, Khanna S (май 2011). «МикроРНК в этиологии и патологии ишемического инсульта». Physiological Genomics . 43 (10): 521–528. doi :10.1152/physiolgenomics.00158.2010. PMC 3110894. PMID 20841499 .
^ Ouyang YB, Stary CM, Yang GY, Giffard R (январь 2013 г.). «микроРНК: инновационные мишени для церебральной ишемии и инсульта». Current Drug Targets . 14 (1): 90–101. doi :10.2174/138945013804806424. PMC 3673881. PMID 23170800 .
^ abc Lewohl JM, Nunez YO, Dodd PR, Tiwari GR, Harris RA, Mayfield RD (ноябрь 2011 г.). "Повышение регуляции микроРНК в мозге людей-алкоголиков". Alcoholism: Clinical and Experimental Research . 35 (11): 1928–37. doi :10.1111/j.1530-0277.2011.01544.x. PMC 3170679. PMID 21651580 .
^ Tapocik JD, Solomon M, Flanigan M, Meinhardt M, Barbier E, Schank JR и др. (июнь 2013 г.). «Скоординированная дисрегуляция мРНК и микроРНК в медиальной префронтальной коре крыс после истории алкогольной зависимости». The Pharmacogenomics Journal . 13 (3): 286–96. doi : 10.1038 /tpj.2012.17. PMC 3546132. PMID 22614244.
^ Gorini G, Nunez YO, Mayfield RD (2013). «Интеграция miRNA и профилирования белков выявляет скоординированные нейроадаптации в мозге мышей, зависимых от алкоголя». PLOS ONE . 8 (12): e82565. Bibcode : 2013PLoSO...882565G. doi : 10.1371/journal.pone.0082565 . PMC 3865091. PMID 24358208 .
^ Tapocik JD, Barbier E, Flanigan M, Solomon M, Pincus A, Pilling A и др. (март 2014 г.). «микроРНК-206 в медиальной префронтальной коре крыс регулирует экспрессию BDNF и употребление алкоголя». The Journal of Neuroscience . 34 (13): 4581–8. doi :10.1523/JNEUROSCI.0445-14.2014. PMC 3965783 . PMID 24672003.
^ Липпай Д., Бала С., Чак Т., Курт-Джонс Е.А., Сабо Г. (2013). «Хроническая вызванная алкоголем микроРНК-155 способствует нейровоспалению в зависимости от TLR4 у мышей». PLOS ONE . 8 (8): e70945. Bibcode :2013PLoSO...870945L. doi : 10.1371/journal.pone.0070945 . PMC 3739772 . PMID 23951048.
^ ab Li J, Li J, Liu X, Qin S, Guan Y, Liu Y и др. (сентябрь 2013 г.). «Профиль экспрессии микроРНК и функциональный анализ показывают, что miR-382 является критическим новым геном алкогольной зависимости». EMBO Molecular Medicine . 5 (9): 1402–14. doi :10.1002/emmm.201201900. PMC 3799494 . PMID 23873704.
^ Romao JM, Jin W, Dodson MV, Hausman GJ, Moore SS, Guan LL (сентябрь 2011 г.). «Регуляция микроРНК в адипогенезе млекопитающих». Experimental Biology and Medicine . 236 (9): 997–1004. doi :10.1258/ebm.2011.011101. PMID 21844119. S2CID 30646787.
^ Skårn M, Namløs HM, Noordhuis P, Wang MY, Meza-Zepeda LA, Myklebost O (апрель 2012 г.). «Адипоцитарная дифференцировка стромальных клеток, полученных из костного мозга человека, модулируется микроРНК-155, микроРНК-221 и микроРНК-222». Стволовые клетки и развитие . 21 (6): 873–83. doi :10.1089/scd.2010.0503. hdl : 10852/40423 . PMID 21756067.
^ Zuo Y, Qiang L, Farmer SR (март 2006 г.). «Активация экспрессии CCAAT/энхансер-связывающего белка (C/EBP) альфа с помощью C/EBP бета во время адипогенеза требует репрессии HDAC1, связанной с активируемым пролифератором пероксисом рецептором-гамма, на промоторе гена C/ebp альфа». Журнал биологической химии . 281 (12): 7960–7. doi : 10.1074/jbc.M510682200 . PMID 16431920.
^ Jun-Hao ET, Gupta RR, Shyh-Chang N (март 2016 г.). «Lin28 и let-7 в метаболической физиологии старения». Тенденции в эндокринологии и метаболизме . 27 (3): 132–141. doi :10.1016/j.tem.2015.12.006. PMID 26811207. S2CID 3614126.
^ Zhu H, Shyh-Chang N, Segrè AV, Shinoda G, Shah SP, Einhorn WS и др. (сентябрь 2011 г.). «Ось Lin28/let-7 регулирует метаболизм глюкозы». Cell . 147 (1): 81–94. doi :10.1016/j.cell.2011.08.033. PMC 3353524 . PMID 21962509.
^ Frost RJ, Olson EN (декабрь 2011 г.). «Контроль гомеостаза глюкозы и чувствительности к инсулину семейством микроРНК Let-7». Труды Национальной академии наук Соединенных Штатов Америки . 108 (52): 21075–80. Bibcode : 2011PNAS..10821075F. doi : 10.1073 /pnas.1118922109 . PMC 3248488. PMID 22160727.
^ Теруэль-Монтойя Р., Розендал Ф.Р., Мартинес С. (февраль 2015 г.). «МикроРНК в гемостазе». Журнал тромбозов и гемостаза . 13 (2): 170–181. дои : 10.1111/jth.12788 . ПМИД 25400249.
^ ab Nourse J, Braun J, Lackner K, Hüttelmaier S, Danckwardt S (ноябрь 2018 г.). «Крупномасштабная идентификация функционального нацеливания микроРНК выявляет кооперативную регуляцию системы гемостаза». Журнал тромбоза и гемостаза . 16 (11): 2233–2245. doi : 10.1111/jth.14290 . PMID 30207063.
^ Nourse J, Danckwardt S (февраль 2021 г.). «Новое обоснование для нацеливания на FXI: выводы из таргета гемостатической микроРНК для новых антикоагулянтных стратегий». Фармакология и терапия . 218 : 107676. doi : 10.1016/j.pharmthera.2020.107676 . PMID 32898547.
^ Berardi E, Pues M, Thorrez L, Sampaolesi M (октябрь 2012 г.). «микроРНК в дифференцировке эмбриональных стволовых клеток». American Journal of Physiology. Heart and Circulatory Physiology . 303 (8): H931–H939. doi :10.1152/ajpheart.00338.2012. PMID 22886416. S2CID 6402014.
^ ab Borchers A, Pieler T (ноябрь 2010 г.). «Программирование плюрипотентных клеток-предшественников, полученных из эмбрионов Xenopus, для создания определенных тканей и органов». Genes . 1 (3): 413–426. doi : 10.3390/ijms232314755 . PMC 9740008 . PMID 36499082.
^ Curaba J, Spriggs A, Taylor J, Li Z, Helliwell C (июль 2012 г.). "Регулирование miRNA в раннем развитии семян ячменя". BMC Plant Biology . 12 (1): 120. doi : 10.1186/1471-2229-12-120 . PMC 3443071. PMID 22838835 .
^ Choudhary A, Kumar A, Kaur H, Kaur N (май 2021 г.). «MiRNA: надсмотрщик растительного мира». Biologia . 76 (5): 1551–1567. Bibcode :2021Biolg..76.1551C. doi :10.1007/s11756-021-00720-1. ISSN 1336-9563. S2CID 233685660.
^ Waheed S, Zeng L (март 2020 г.). «Критическая роль микроРНК в регуляции времени цветения и развития цветка». Genes . 11 (3): 319. doi : 10.3390/genes11030319 . PMC 7140873 . PMID 32192095.
^ Wong CE, Zhao YT, Wang XJ, Croft L, Wang ZH, Haerizadeh F, et al. (Май 2011). «МикроРНК в апикальной меристеме побега сои». Журнал экспериментальной ботаники . 62 (8): 2495–2506. doi :10.1093/jxb/erq437. hdl : 10536/DRO/DU:30106047 . PMID 21504877.
^ Куреши А, Такур Н, Монга И, Такур А, Кумар М (1 января 2014 г.). «VIRmiRNA: всеобъемлющий ресурс для экспериментально подтвержденных вирусных miRNA и их целей». База данных . 2014 : bau103. doi :10.1093/database/bau103. PMC 4224276. PMID 25380780 .
^ Кумар М. "VIRmiRNA". Ресурс по экспериментальным вирусным miRNA и их целям . Центр биоинформатики, CSIR-IMTECH.
^ Tuddenham L, Jung JS, Chane-Woon-Ming B, Dölken L, Pfeffer S (февраль 2012 г.). «Глубокое секвенирование малых РНК выявляет микроРНК и другие малые некодирующие РНК из вируса герпеса человека 6B». Журнал вирусологии . 86 (3): 1638–49. doi :10.1128/JVI.05911-11. PMC 3264354. PMID 22114334 .
^ Zheng H, Fu R, Wang JT, Liu Q, Chen H, Jiang SW (апрель 2013 г.). «Достижения в методах прогнозирования целей микроРНК». International Journal of Molecular Sciences . 14 (4): 8179–87. doi : 10.3390/ijms14048179 . PMC 3645737. PMID 23591837 .
^ Агарвал В., Белл Г. В., Нам Дж. В., Бартель Д. П. (август 2015 г.). «Прогнозирование эффективных целевых участков микроРНК в мРНК млекопитающих». eLife . 4 : e05005. doi : 10.7554/eLife.05005 . PMC 4532895 . PMID 26267216.
Дальнейшее чтение
Определение и классификация miRNA: Ambros V, Bartel B, Bartel DP, Burge CB, Carrington JC, Chen X и др. (март 2003 г.). «Единая система аннотации микроРНК». RNA . 9 (3): 277–9. doi :10.1261/rna.2183803. PMC 1370393 . PMID 12592000.
Научный обзор малых РНК: Baulcombe D (сентябрь 2002 г.). «ДНК-события. Микрокосм РНК». Science . 297 (5589): 2002–3. doi :10.1126/science.1077906. PMID 12242426. S2CID 82531727.
Открытие lin-4 , первой обнаруженной miRNA: Lee RC, Feinbaum RL, Ambros V (декабрь 1993 г.). «Гетерохронный ген C. elegans lin-4 кодирует малые РНК с антисмысловой комплементарностью к lin-14». Cell . 75 (5): 843–54. doi : 10.1016/0092-8674(93)90529-Y . PMID 8252621.
Внешние ссылки
Найдите термин «микроРНК» или «миРНК» в Викисловаре, бесплатном словаре.
База данных miRBase
miRTarBase — экспериментально подтвержденная база данных взаимодействий микроРНК с мишенями.
semirna, веб-приложение для поиска микроРНК в геноме растений.
ONCO.IO: Интегративный ресурс для анализа микроРНК и факторов транскрипции при раке.
MirOB Архивировано 4 марта 2014 г. на Wayback Machine : MicroRNA нацелена на базу данных, анализ данных и инструмент визуализации данных.
База данных ChIPBase: База данных открытого доступа для расшифровки факторов транскрипции , которые участвовали или влияли на транскрипцию микроРНК из данных ChIP-seq.
Анимированное видео процесса биогенеза микроРНК.
Реагенты для модуляции микроРНК, позволяющие повышать или подавлять функцию эндогенной зрелой микроРНК
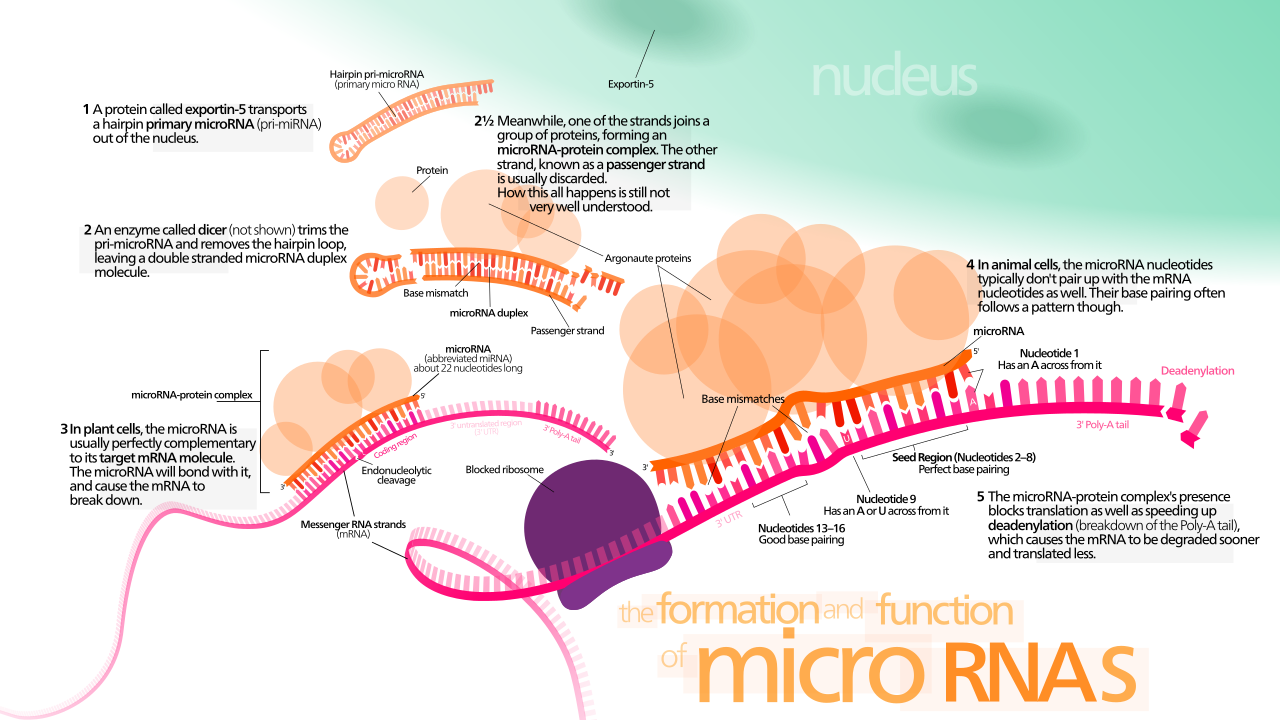







{kind=link}