Биология развития – это изучение процесса роста и развития животных и растений . Биология развития также охватывает биологию регенерации , бесполого размножения , метаморфоза , а также роста и дифференциации стволовых клеток во взрослом организме.
Основными процессами, участвующими в эмбриональном развитии животных, являются: формирование паттерна ткани (посредством региональной спецификации и дифференцировки клеток по паттерну ); рост тканей ; и морфогенез тканей .
В развитии растений происходят те же процессы, что и у животных. Однако растительные клетки в основном неподвижны, поэтому морфогенез достигается за счет дифференциального роста без движений клеток. Кроме того, индуктивные сигналы и задействованные гены отличаются от тех, которые контролируют развитие животных.

Дифференциация клеток — это процесс, в результате которого в развитии возникают различные функциональные типы клеток. Например, нейроны, мышечные волокна и гепатоциты (клетки печени) являются хорошо известными типами дифференцированных клеток. Дифференцированные клетки обычно производят большое количество нескольких белков, необходимых для выполнения их специфической функции, и это придает им характерный внешний вид, позволяющий распознавать их под световым микроскопом. Гены, кодирующие эти белки, высокоактивны. Обычно их структура хроматина очень открыта, что обеспечивает доступ ферментам транскрипции, а специфические факторы транскрипции связываются с регуляторными последовательностями ДНК, чтобы активировать экспрессию генов. [1] [2] Например, NeuroD является ключевым фактором транскрипции для дифференцировки нейронов, миогенин для дифференцировки мышц и HNF4 для дифференцировки гепатоцитов. Дифференциация клеток обычно является заключительной стадией развития, которой предшествуют несколько состояний детерминации, которые визуально не дифференцируются. Одна ткань, образованная из одного типа клеток-предшественников или стволовых клеток, часто состоит из нескольких дифференцированных типов клеток. Контроль их образования включает процесс латерального ингибирования [3] , основанный на свойствах сигнального пути Notch . [4] Например, в нервной пластинке эмбриона эта система генерирует популяцию нейрональных клеток-предшественников, в которых высока экспрессия NeuroD.
Регенерация указывает на способность восстановить недостающую часть. [5] Это очень распространено среди растений, которые демонстрируют непрерывный рост, а также среди колониальных животных, таких как гидроиды и асцидии. Но наибольший интерес биологи развития проявляют к регенерации частей свободноживущих животных. В частности, четыре модели стали предметом многочисленных исследований. Двое из них обладают способностью регенерировать целые тела: гидра , которая может регенерировать любую часть полипа из небольшого фрагмента, [6] и планарийные черви, которые обычно могут регенерировать как голову, так и хвост. [7] В обоих этих примерах наблюдается непрерывный клеточный оборот, питаемый стволовыми клетками , и, по крайней мере, у планарий, по крайней мере, некоторые из стволовых клеток оказались плюрипотентными . [8] Две другие модели показывают только дистальную регенерацию придатков. Это придатки насекомых, обычно ноги полуметаболических насекомых, таких как сверчок [9] и конечности хвостатых амфибий . [10] В настоящее время доступен значительный объем информации о регенерации конечностей амфибий, и известно, что каждый тип клеток регенерирует сам, за исключением соединительных тканей, где происходит значительная взаимоконверсия между хрящом, дермой и сухожилиями. Что касается структуры структур, то это контролируется повторной активацией сигналов, активных в эмбрионе. До сих пор ведутся споры по поводу старого вопроса о том, является ли регенерация «первозданным» или «адаптивным» свойством. [11] Если это так, то с улучшением знаний мы могли бы ожидать, что сможем улучшить регенеративные способности у людей. Если последнее, то предполагается, что каждый случай регенерации возник в результате естественного отбора в обстоятельствах, специфичных для данного вида, поэтому не следует ожидать каких-либо общих правил.

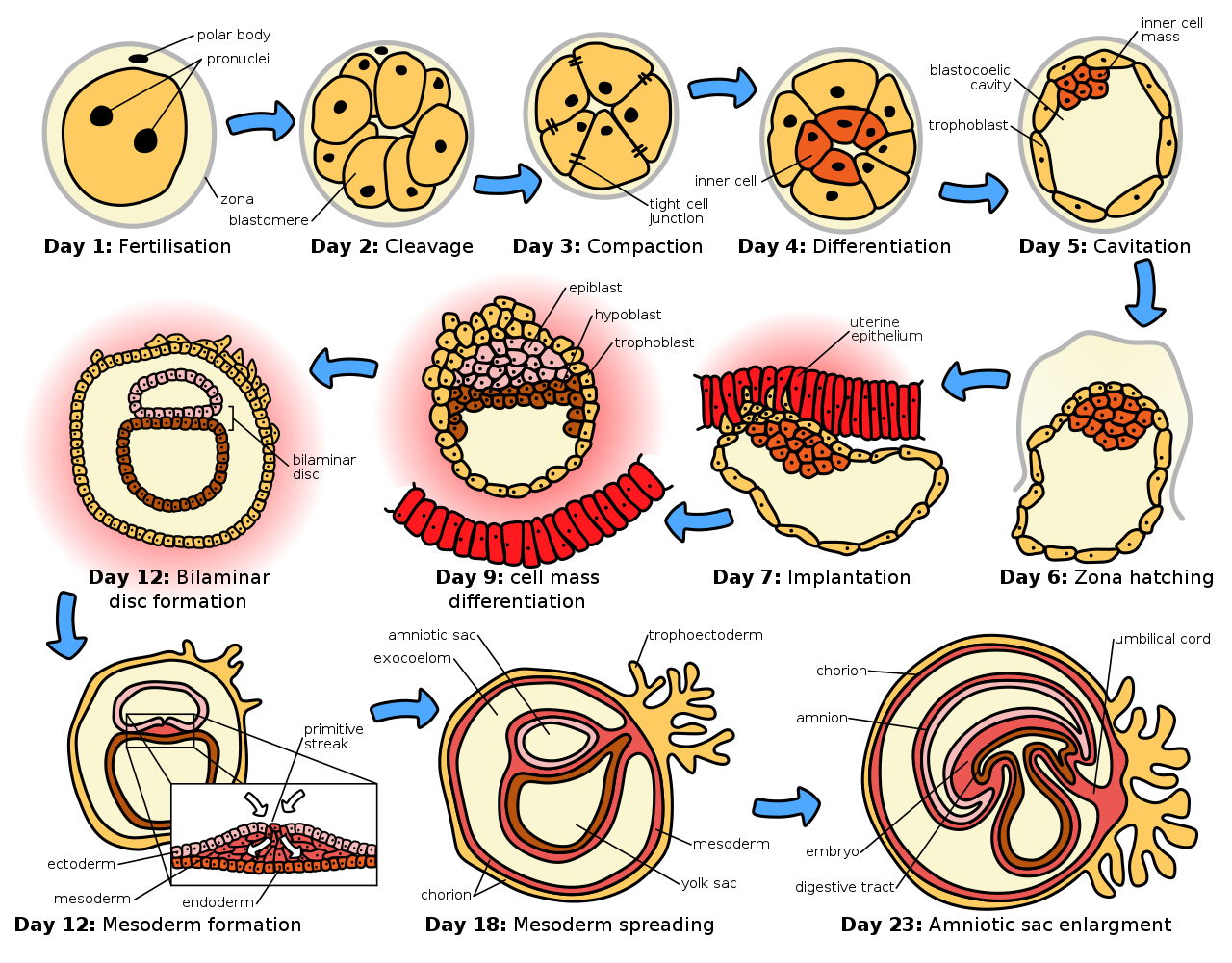
Сперматозоид и яйцеклетка сливаются в процессе оплодотворения, образуя оплодотворенную яйцеклетку или зиготу . [12] Она проходит период делений с образованием шара или листа одинаковых клеток, называемых бластулой или бластодермой . Эти клеточные деления обычно происходят быстро и без роста, поэтому дочерние клетки составляют половину размера материнской клетки, а весь эмбрион остается примерно того же размера. Их называют делениями расщепления .
Первичные зародышевые клетки эпибласта мыши (см. рисунок: «Начальные стадии эмбриогенеза человека ») подвергаются обширному эпигенетическому перепрограммированию. [13] Этот процесс включает в себя деметилирование ДНК по всему геному , реорганизацию хроматина и стирание эпигенетического импринта, что приводит к тотипотентности . [13] Деметилирование ДНК осуществляется с помощью процесса, в котором используется путь эксцизионной репарации оснований ДНК . [14]
Морфогенетические движения преобразуют клеточную массу в трехслойную структуру, состоящую из многоклеточных листов, называемых эктодермой , мезодермой и энтодермой . Эти листы известны как зародышевые листки . Это процесс гаструляции . Во время дробления и гаструляции происходят первые события региональной спецификации. Помимо формирования самих трех зародышевых листков, они часто образуют внеэмбриональные структуры, такие как плацента млекопитающих , необходимые для поддержки и питания эмбриона [15] , а также устанавливают различия в детерминации вдоль переднезадней оси (голова, туловище). и хвост). [16]
Региональная спецификация инициируется наличием цитоплазматических детерминант в одной части зиготы. Клетки, содержащие детерминанту, становятся сигнальным центром и выделяют индуцирующий фактор. Поскольку индуцирующий фактор вырабатывается в одном месте, диффундирует и распадается, он образует градиент концентрации, высокий вблизи клеток-источников и низкий вдали от них. [17] [18] Остальные клетки эмбриона, которые не содержат детерминанту, способны реагировать на различные концентрации путем активации специфических генов контроля развития. В результате создается ряд зон, расположенных на все большем расстоянии от сигнального центра. В каждой зоне активируется различная комбинация генов контроля развития. [19] Эти гены кодируют факторы транскрипции , которые активируют новые комбинации активности генов в каждой области. Помимо других функций, эти факторы транскрипции контролируют экспрессию генов, придающих специфические адгезивные и подвижные свойства клеткам, в которых они активны. Из-за этих различных морфогенетических свойств клетки каждого зародышевого листка перемещаются, образуя листы, так что эктодерма оказывается снаружи, мезодерма — посередине, а энтодерма — внутри. [20] [21]

Морфогенетические движения не только изменяют форму и структуру эмбриона, но, приводя клеточные листы в новые пространственные взаимоотношения, они также делают возможными новые фазы передачи сигналов и реакции между ними. Кроме того, первые морфогенетические движения эмбриогенеза, такие как гаструляция, эпиболия и скручивание , напрямую активируют пути, участвующие в спецификации эндомезодермы посредством процессов механотрансдукции. [22] [23] Было высказано предположение, что это свойство эволюционно унаследовано от спецификации эндомезодермы, механически стимулируемой гидродинамическим потоком морской среды у первых животных организмов (первых многоклеточных животных). [24] Скручивание вдоль оси тела за счет левосторонней хиральности встречается у всех хордовых (включая позвоночных) и рассматривается в теории осевого скручивания . [25]
Рост эмбрионов в основном автономен. [26] Для каждой территории клеток скорость роста контролируется комбинацией активных генов. Свободноживущие эмбрионы не растут в массе, поскольку у них нет внешнего источника питания. Но эмбрионы, питаемые плацентой или внеэмбриональным желтком, могут расти очень быстро, и изменения относительной скорости роста между частями этих организмов помогают создать окончательную общую анатомию.
Весь процесс нужно согласовать во времени и как это контролируется непонятно. Могут существовать главные часы, способные взаимодействовать со всеми частями эмбриона и контролировать ход событий, или время может зависеть просто от локальных причинных последовательностей событий. [27]
Процессы развития очень очевидны в процессе метаморфоза . Это происходит у различных видов животных. Хорошо известные примеры можно увидеть у лягушек, которые обычно вылупляются головастиками и превращаются во взрослую лягушку, а также у некоторых насекомых, которые вылупляются как личинки, а затем на стадии куколки перестраиваются во взрослую форму.
Все перечисленные выше процессы развития происходят в ходе метаморфоза. Примеры, которые были особенно хорошо изучены, включают потерю хвоста и другие изменения в головастике лягушки Xenopus , [28] [29] и биологию имагинальных дисков, из которых образуются части тела взрослой мухи Drosophila melanogaster . [30] [31]
Развитие растений — это процесс, в ходе которого структуры возникают и созревают по мере роста растения. Он изучается в анатомии и физиологии растений , а также в морфологии растений.
Растения на протяжении всей своей жизни постоянно производят новые ткани и структуры из меристем [32] , расположенных на кончиках органов или между зрелыми тканями. Таким образом, живое растение всегда имеет эмбриональные ткани. Напротив, эмбрион животного очень рано производит все части тела, которые у него когда-либо будут в жизни. Когда животное рождается (или вылупляется из яйца), у него есть все части тела, и с этого момента оно становится только крупнее и взрослее.
Свойства организации, наблюдаемые в растении, — это эмерджентные свойства , которые представляют собой нечто большее, чем сумма отдельных частей. «Сборка этих тканей и функций в единый многоклеточный организм дает не только характеристики отдельных частей и процессов, но и совершенно новый набор характеристик, который невозможно было бы предсказать на основе изучения отдельных частей». [33]
Сосудистое растение начинается из одноклеточной зиготы , образующейся в результате оплодотворения яйцеклетки сперматозоидом. С этого момента оно начинает делиться, образуя зародыш растения в процессе эмбриогенеза . При этом образующиеся клетки организуются так, что один конец становится первым корнем, а другой конец образует верхушку побега. У семенных растений у зародыша разовьется один или несколько «семенных листьев» ( семядолей ). К концу эмбриогенеза молодое растение будет иметь все части, необходимые для начала жизни.
Как только эмбрион прорастает из семени или родительского растения, он начинает производить дополнительные органы (листья, стебли и корни) в процессе органогенеза . Новые корни вырастают из корневых меристем , расположенных на кончике корня, а новые стебли и листья вырастают из меристем побега , расположенных на кончике побега. [34] Ветвление происходит, когда небольшие скопления клеток, оставшиеся от меристемы и еще не подвергшиеся клеточной дифференцировке с образованием специализированной ткани, начинают расти как верхушка нового корня или побега. Рост любой такой меристемы на кончике корня или побега называется первичным ростом и приводит к удлинению этого корня или побега. Вторичный рост приводит к расширению корня или побега за счет деления клеток камбия . [35]
Помимо роста за счет деления клеток , растение может расти за счет удлинения клеток . [36] Это происходит, когда отдельные клетки или группы клеток становятся длиннее. Не все растительные клетки вырастают до одинаковой длины. Когда клетки на одной стороне стебля растут длиннее и быстрее, чем клетки на другой стороне, в результате стебель изгибается в сторону более медленно растущих клеток. Этот направленный рост может происходить через реакцию растения на определенный раздражитель, такой как свет ( фототропизм ), гравитация ( гравитропизм ), вода ( гидротропизм ) и физический контакт ( тигмотропизм ).
Рост и развитие растений опосредованы специфическими фитогормонами и регуляторами роста растений (РГР) (Росс и др., 1983). [37] На уровень эндогенных гормонов влияют возраст растения, холодостойкость, период покоя и другие метаболические условия; фотопериод, засуха, температура и другие внешние условия среды; и экзогенные источники ГРР, например, применяемые извне и ризосферного происхождения.
Растения демонстрируют естественные различия в своей форме и строении. Хотя все организмы варьируются от человека к человеку, растения демонстрируют дополнительный тип вариаций. Внутри одной особи повторяются части, которые по форме и строению могут отличаться от других подобных частей. Эту вариацию легче всего увидеть на листьях растения, хотя и в других органах, таких как стебли и цветы, могут наблюдаться аналогичные вариации. Есть три основные причины этого изменения: позиционные эффекты, воздействие окружающей среды и молодость.
Факторы транскрипции и сети регуляции транскрипции играют ключевую роль в морфогенезе растений и их эволюции. Во время посадки растений появилось множество новых семейств транскрипционных факторов, которые преимущественно встраиваются в сети многоклеточного развития, размножения и развития органов, способствуя более сложному морфогенезу наземных растений. [38]
Большинство наземных растений имеют общего предка — многоклеточные водоросли. Пример эволюции морфологии растений можно увидеть у харофитов. Исследования показали, что харофиты обладают признаками, гомологичными наземным растениям. Существуют две основные теории эволюции морфологии растений: гомологичная теория и антитетическая теория. Общепринятой теорией эволюции морфологии растений является антитетическая теория. Противоположная теория утверждает, что множественные митотические деления, происходящие перед мейозом, вызывают развитие спорофита. Тогда спорофит будет развиваться как самостоятельный организм. [39]
Большая часть исследований в области биологии развития в последние десятилетия была сосредоточена на использовании небольшого числа модельных организмов . Оказалось, что в животном мире механизмы развития во многом консервативны. На раннем этапе развития все виды позвоночных используют по существу одни и те же индуктивные сигналы и одни и те же гены, кодирующие региональную идентичность. Даже беспозвоночные используют схожий набор сигналов и генов, хотя сформированные части тела существенно различаются. Каждый из модельных организмов обладает определенными экспериментальными преимуществами, которые позволили им стать популярными среди исследователей. В одном смысле они являются «моделями» всего животного мира, а в другом — «моделями» человеческого развития, которое трудно изучать непосредственно как по этическим, так и по практическим причинам. Модельные организмы оказались наиболее полезными для выяснения широкой природы механизмов развития. Чем больше деталей ищется, тем больше они отличаются друг от друга и от человека.
Также популярными для некоторых целей были морские ежи [48] [40] и асцидии . [49] Для изучения регенерации хвостатых амфибий используются такие как аксолотль Ambystoma mexicanum , [50] а также планарийные черви, такие как Schmidtea mediterranea . [7] Органоиды также были продемонстрированы как эффективная модель развития. [51] Развитие растений было сосредоточено на кресс-салате Arabidopsis thaliana в качестве модельного организма. [52]
{{cite book}}: CS1 maint: location missing publisher (link){{cite book}}: |journal=игнорируется ( помощь )