Слово haem происходит от греческого αἷμα haima — «кровь».
Модель заполнения пространства субъединицы Fe- протопорфирина IX гема B. Аксиальные лиганды опущены. Цветовая схема: серый = железо, синий = азот, черный = углерод, белый = водород, красный = кислород
Гемопротеины выполняют разнообразные биологические функции, включая транспортировку двухатомных газов, химический катализ , обнаружение двухатомных газов и перенос электронов . Гемовое железо служит источником или поглотителем электронов во время переноса электронов или окислительно-восстановительной химии. В пероксидазных реакциях молекула порфирина также служит источником электронов, будучи способной делокализировать радикальные электроны в сопряженном кольце. При транспортировке или обнаружении двухатомных газов газ связывается с гемовым железом. Во время обнаружения двухатомных газов связывание газового лиганда с гемовым железом вызывает конформационные изменения в окружающем белке. [10] В целом, двухатомные газы связываются только с восстановленным гемом, как двухвалентное железо Fe(II), в то время как большинство пероксидаз циклируют между Fe(III) и Fe(IV), а гемовые протеины участвуют в митохондриальном окислительно-восстановительном цикле между Fe(II) и Fe(III).
Было высказано предположение, что первоначальной эволюционной функцией гемопротеинов был перенос электронов в примитивных путях фотосинтеза на основе серы у предковых цианобактерий -подобных организмов до появления молекулярного кислорода . [11]
Гемопротеины достигают своего замечательного функционального разнообразия путем изменения среды макроцикла гема в белковой матрице. [12] Например, способность гемоглобина эффективно доставлять кислород тканям обусловлена специфическими аминокислотными остатками, расположенными вблизи молекулы гема. [13] Гемоглобин обратимо связывается с кислородом в легких, когда pH высокий , а концентрация углекислого газа низкая. Когда ситуация обратная (низкий pH и высокая концентрация углекислого газа), гемоглобин будет выделять кислород в ткани. Это явление, которое гласит, что сродство гемоглобина к связыванию кислорода обратно пропорционально как кислотности , так и концентрации углекислого газа, известно как эффект Бора . [14] Молекулярный механизм , лежащий в основе этого эффекта, заключается в стерической организации цепи глобина ; остаток гистидина , расположенный рядом с группой гема, становится положительно заряженным в кислых условиях (которые вызваны растворенным CO2 в работающих мышцах и т. д.), высвобождая кислород из группы гема. [15]
Типы
Основные гемы
Существует несколько биологически важных видов гема:
Структура Fe-порфириновой субъединицы гема B.Структура Fe-порфириновой субъединицы гема А. [16] Гем А синтезируется из гема В. В двух последовательных реакциях 17-гидроксиэтилфарнезильный фрагмент добавляется в положение 2, а альдегид добавляется в положение 8. [17]
Наиболее распространенным типом является гем B ; другие важные типы включают гем A и гем C. Изолированные гемы обычно обозначаются заглавными буквами, тогда как гемы, связанные с белками, обозначаются строчными буквами. Цитохром a относится к гему A в определенной комбинации с мембранным белком, образующим часть цитохром c оксидазы . [18]
Другие гемы
Следующая система нумерации атомов углерода порфиринов является более старой системой, используемой биохимиками, а не системой нумерации 1–24, рекомендованной ИЮПАК , которая показана в таблице выше.
Гем l является производным гема B, который ковалентно связан с белком лактопероксидазы , эозинофильной пероксидазы и тиреоидной пероксидазы . Добавление пероксида с глутамил -375 и аспартил -225 лактопероксидазы образует сложноэфирные связи между этими аминокислотными остатками и гемовыми 1- и 5-метильными группами соответственно. [19] Считается, что подобные сложноэфирные связи с этими двумя метильными группами образуются в эозинофильных и тиреоидных пероксидазах. Гем l является одной из важных характеристик животных пероксидаз; растительные пероксидазы включают гем B. Лактопероксидаза и эозинофильная пероксидаза являются защитными ферментами, ответственными за уничтожение вторгающихся бактерий и вирусов. Тиреоидная пероксидаза является ферментом, катализирующим биосинтез важных гормонов щитовидной железы. Поскольку лактопероксидаза уничтожает вторгающиеся организмы в легких и экскрементах, она считается важным защитным ферментом. [20]
Гем m является производным гема B, ковалентно связанным в активном центре миелопероксидазы . Гем m содержит две эфирные связи в гемовых 1- и 5-метильных группах, также присутствующих в геме l других пероксидаз млекопитающих, таких как лактопероксидаза и эозинофильная пероксидаза. Кроме того, образуется уникальная сульфонамидная ионная связь между серой остатка метиониламинокислоты и гемовой 2-винильной группой, что дает этому ферменту уникальную способность легко окислять ионы хлорида и бромида до гипохлорита и гипобромита. Миелопероксидаза присутствует в нейтрофилах млекопитающих и отвечает за уничтожение вторгающихся бактерий и вирусных агентов. Возможно, она синтезирует гипобромит по «ошибке». И гипохлорит, и гипобромит являются очень реактивными видами, ответственными за производство галогенированных нуклеозидов, которые являются мутагенными соединениями. [21] [22]
Гем D — это еще одно производное гема B, но в котором боковая цепь пропионовой кислоты у углерода в положении 6, которая также гидроксилирована, образует γ- спиролактон . Кольцо III также гидроксилировано в положении 5 в конформации транс к новой лактонной группе. [23] Гем D — это место восстановления кислорода до воды у многих типов бактерий при низком напряжении кислорода. [24]
Гем S связан с гемом B тем, что имеет формильную группу в позиции 2 вместо 2-винильной группы. Гем S обнаружен в гемоглобине нескольких видов морских червей. Правильные структуры гема B и гема S были впервые выяснены немецким химиком Гансом Фишером . [25]
Названия цитохромов обычно (но не всегда) отражают типы содержащихся в них гемов: цитохром а содержит гем А, цитохром с содержит гем С и т. д. Эта условность, возможно, впервые была введена с публикацией структуры гема А.
Использование заглавных букв для обозначения типа гема
Практика обозначения гемов заглавными буквами была формализована в сноске в статье Пуустинена и Викстрома [26] , где объясняется, при каких условиях следует использовать заглавную букву: «мы предпочитаем использовать заглавные буквы для описания структуры гема как изолированной. Строчные буквы затем могут свободно использоваться для цитохромов и ферментов, а также для описания отдельных групп гемов, связанных с белками (например, комплексы цитохрома bc и aa3, цитохром b 5 , гем c 1 комплекса bc 1 , гем a 3 комплекса aa 3 и т. д.)». Другими словами, химическое соединение будет обозначено заглавной буквой, но конкретные примеры в структурах — строчными. Таким образом, цитохромоксидаза, которая имеет в своей структуре два гема A (гем a и гем a 3 ), содержит два моля гема A на моль белка. Цитохром bc 1 , с гемами b H , b L , и c 1 , содержит гем B и гем C в соотношении 2:1. Похоже, что эта практика возникла в статье Коги и Йорка, в которой продукт новой процедуры выделения гема цитохрома aa3 был обозначен как гем A, чтобы отличить его от предыдущих препаратов: «Наш продукт не во всех отношениях идентичен гемому a, полученному в растворе другими исследователями путем восстановления гемина a, выделенного ранее (2). По этой причине мы будем обозначать наш продукт как гем A до тех пор, пока не будут объяснены очевидные различия». [27] В более поздней статье [28] группа Коги использует заглавные буквы для выделенных гемов B и C, а также A.
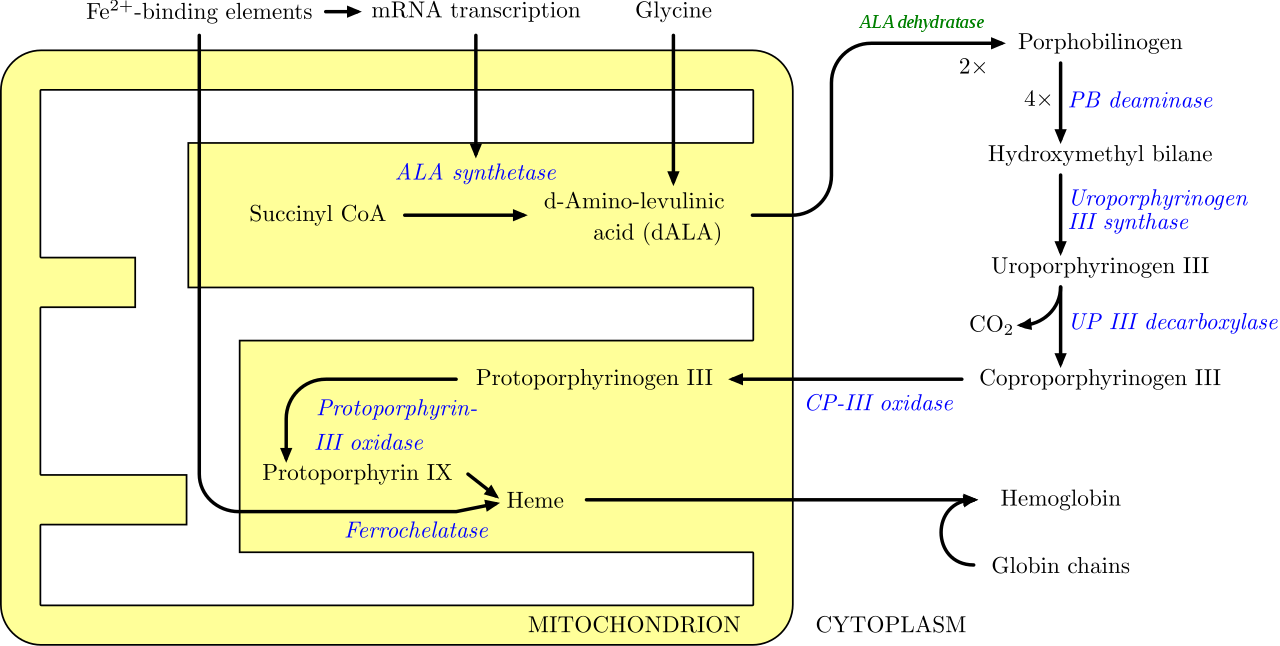
Ферментативный процесс, который производит гем, правильно называется синтезом порфирина , так как все промежуточные продукты являются тетрапирролами , которые химически классифицируются как порфирины. Процесс высоко консервативен в биологии. У людей этот путь служит почти исключительно для образования гема. У бактерий он также производит более сложные вещества, такие как кофактор F430 и кобаламин ( витамин B 12 ). [29]
Путь инициируется синтезом δ-аминолевулиновой кислоты (dALA или δALA) из аминокислоты глицина и сукцинил-КоА из цикла лимонной кислоты (цикла Кребса). Скорость-лимитирующий фермент, ответственный за эту реакцию, ALA-синтаза , отрицательно регулируется концентрацией глюкозы и гема. Механизм ингибирования ALA гемом или гемином заключается в снижении стабильности синтеза мРНК и снижении поступления мРНК в митохондрии. Этот механизм имеет терапевтическое значение: инфузия гем-аргината или гематина и глюкозы может прекратить приступы острой перемежающейся порфирии у пациентов с врожденным нарушением метаболизма этого процесса, за счет снижения транскрипции ALA-синтазы. [30]
Органами, в основном участвующими в синтезе гема, являются печень (в которой скорость синтеза сильно варьируется в зависимости от системного пула гема) и костный мозг (в котором скорость синтеза гема относительно постоянна и зависит от выработки глобиновой цепи), хотя каждой клетке для правильного функционирования требуется гем. Однако из-за его токсичных свойств такие белки, как эмопексин (Hx), необходимы для поддержания физиологических запасов железа, чтобы они могли использоваться в синтезе. [31] Гем рассматривается как промежуточная молекула в катаболизме гемоглобина в процессе метаболизма билирубина . Дефекты различных ферментов в синтезе гема могут привести к группе расстройств, называемых порфириями, которые включают острую перемежающуюся порфирию , врожденную эритропоэтическую порфирию , позднюю кожную порфирию , наследственную копропорфирию , смешанную порфирию и эритропоэтическую протопорфирию . [32]
Синтез для еды
Impossible Foods , производители заменителей мяса на растительной основе , используют ускоренный процесс синтеза гема с участием леггемоглобина корня сои и дрожжей , добавляя полученный гем в такие продукты, как постные ( веганские ) котлеты для бургеров Impossible. ДНК для производства леггемоглобина была извлечена из клубеньков корня сои и экспрессирована в дрожжевых клетках для перепроизводства гема для использования в постных бургерах. [33] Этот процесс, как утверждается, создает мясной вкус в полученных продуктах. [34] [35]
Деградация
Распад гема
Деградация начинается внутри макрофагов селезенки , которые удаляют старые и поврежденные эритроциты из кровообращения.
На первом этапе гем преобразуется в биливердин ферментом гемоксигеназой (HO). [36] НАДФН используется в качестве восстановителя, молекулярный кислород вступает в реакцию, образуется оксид углерода (CO), а железо высвобождается из молекулы в виде иона железа (Fe2 + ). [37] CO действует как клеточный мессенджер и выполняет функцию вазодилатации. [38]
Кроме того, деградация гема, по-видимому, является эволюционно-консервативной реакцией на окислительный стресс . Вкратце, когда клетки подвергаются воздействию свободных радикалов , происходит быстрая индукция экспрессии стресс-чувствительного изофермента гем-оксигеназы-1 (HMOX1), который катаболизирует гем (см. ниже). [39] Причина, по которой клетки должны экспоненциально увеличивать свою способность деградировать гем в ответ на окислительный стресс, остается неясной, но это, по-видимому, является частью цитопротекторного ответа, который позволяет избежать пагубного воздействия свободного гема. Когда накапливается большое количество свободного гема, системы детоксикации/деградации гема перегружаются, что позволяет гему оказывать свое разрушительное воздействие. [31]
Эта форма билирубина выводится из печени в желчь . Выделение билирубина из печени в желчные канальцы является активным, энергозависимым и лимитирующим скорость процессом. Кишечные бактерии деконъюгируют диглюкуронид билирубина, высвобождая свободный билирубин, который может быть либо реабсорбирован, либо восстановлен до уробилиногена бактериальным ферментом билирубинредуктазой. [42]
Часть уробилиногена поглощается клетками кишечника и транспортируется в почки и выводится с мочой ( уробилин , который является продуктом окисления уробилиногена и отвечает за желтый цвет мочи). Остальная часть перемещается вниз по пищеварительному тракту и преобразуется в стеркобилиноген . Он окисляется до стеркобилина , который выводится и отвечает за коричневый цвет кала . [43]
В здоровье и болезни
При гомеостазе реактивность гема контролируется его вставкой в «гемовые карманы» гемопротеинов. [ необходима цитата ] Однако при окислительном стрессе некоторые гемопротеины, например гемоглобин, могут высвобождать свои простетические группы гема. [44] [45] Несвязанный с белком (свободный) гем, полученный таким образом, становится высокоцитотоксичным, скорее всего, из-за атома железа, содержащегося в его кольце протопорфирина IX, который может действовать как реагент Фентона , чтобы беспрепятственно катализировать производство свободных радикалов. [46] Он катализирует окисление и агрегацию белка, образование цитотоксического липидного пероксида посредством липидного перекисного окисления и повреждает ДНК посредством окислительного стресса. Из-за своих липофильных свойств он повреждает липидные бислои в органеллах, таких как митохондрии и ядра. [47] Эти свойства свободного гема могут сенсибилизировать различные типы клеток, чтобы они подвергались запрограммированной клеточной смерти в ответ на провоспалительные агонисты, вредный эффект, который играет важную роль в патогенезе некоторых воспалительных заболеваний, таких как малярия [48] и сепсис . [49]
Рак
Существует связь между высоким потреблением гемового железа, полученного из мяса, и повышенным риском колоректального рака . [50]
Американский институт исследований рака (AICR) и Всемирный фонд исследований рака (WCRF) в своем отчете за 2018 год пришли к выводу, что имеются ограниченные, но убедительные доказательства того, что продукты, содержащие гемовое железо, повышают риск колоректального рака. [51] Обзор 2019 года показал, что потребление гемового железа связано с повышенным риском рака молочной железы . [52]
Гены
Следующие гены являются частью химического пути образования гема:
ALAD : аминолевулиновая кислота, δ- дегидратаза (дефицит вызывает порфирию с дефицитом аладегидратазы) [53]
^ Hodgson E, Roe RM, Mailman RB, Chambers JE, ред. (2015). "H". Словарь токсикологии (3-е изд.). Academic Press. стр. 173–184. doi :10.1016/B978-0-12-420169-9.00008-4. ISBN 978-0-12-420169-9. Получено 21.02.2024 .
^ Bloomer JR (1998). «Печеночный метаболизм порфиринов и гема». Журнал гастроэнтерологии и гепатологии . 13 (3): 324–329. doi : 10.1111/j.1440-1746.1998.01548.x . PMID 9570250. S2CID 25224821.
^ Химия IU (2009). "Гемы (производные гема)". Сборник химических терминов ИЮПАК . ИЮПАК. doi :10.1351/goldbook.H02773. ISBN978-0-9678550-9-7. Архивировано из оригинала 22 августа 2017 . Получено 28 апреля 2018 .
^ Стандартный текст по биохимии определяет гем как «железо-порфириновую простетическую группу гемовых белков» (Нельсон, Д.Л.; Кокс, М.М. «Ленингер, Принципы биохимии», 3-е изд., Уорт Паблишинг: Нью-Йорк, 2000. ISBN 1-57259-153-6 .)
^ Poulos TL (2014-04-09). «Структура и функции гем-фермента». Chemical Reviews . 114 (7): 3919–3962. doi :10.1021/cr400415k. ISSN 0009-2665. PMC 3981943. PMID 24400737 .
^ Paoli M (2002). «Структурно-функциональные связи в гем-протеинах» (PDF) . DNA Cell Biol . 21 (4): 271–280. doi :10.1089/104454902753759690. hdl :20.500.11820/67200894-eb9f-47a2-9542-02877d41fdd7. PMID 12042067. S2CID 12806393. Архивировано (PDF) из оригинала 24.07.2018.
^ Alderton W (2001). «Синтазы оксида азота: структура, функция и ингибирование». Biochem. J . 357 (3): 593–615. doi :10.1042/bj3570593. PMC 1221991 . PMID 11463332.
^ Milani M (2005). «Структурные основы связывания гема и распознавания двухатомных лигандов в укороченных гемоглобинах». J. Inorg. Biochem . 99 (1): 97–109. doi :10.1016/j.jinorgbio.2004.10.035. PMID 15598494.
^ Хардисон Р. (1999). «Эволюция гемоглобина: исследования очень древнего белка показывают, что изменения в регуляции генов являются важной частью эволюционной истории». American Scientist . 87 (2): 126. doi :10.1511/1999.20.809. S2CID 123532036.
^ Poulos T (2014). «Структура и функция гем-фермента». Chem. Rev. 114 ( 7): 3919–3962. doi :10.1021/cr400415k. PMC 3981943. PMID 24400737 .
^ Том CS (2013). «Варианты гемоглобина: биохимические свойства и клинические корреляты». Cold Spring Harbor Perspectives in Medicine . 3 (3): a011858. doi :10.1101/cshperspect.a011858. PMC 3579210. PMID 23388674 .
^ Бор, Хассельбальх, Крог. «О биологически важной связи — влиянии содержания углекислого газа в крови на ее связывание кислорода». Архивировано из оригинала 18.04.2017.{{cite journal}}: Цитировать журнал требует |journal=( помощь )
^ Ackers GK, Holt JM (2006). «Асимметричная кооперативность в симметричном тетрамере: гемоглобин человека». J. Biol. Chem . 281 (17): 11441–3. doi : 10.1074/jbc.r500019200 . PMID 16423822. S2CID 6696041.
^ Caughey WS, Smythe GE, O'Keeffe DH, Maskasky JE, Smith ML (1975). «Гем A цитохром c оксидазы: структура и свойства: сравнение с гемами B, C и S и их производными». J. Biol. Chem . 250 (19): 7602–7622. doi : 10.1016/S0021-9258(19)40860-0 . PMID 170266.
^ Hegg EL (2004). «Гем А-синтаза не включает молекулярный кислород в формильную группу гема А». Биохимия . 43 (27): 8616–8624. doi :10.1021/bi049056m. PMID 15236569.
^ Yoshikawa S (2012). «Структурные исследования цитохром с оксидазы бычьего сердца». Biochim. Biophys. Acta . 1817 (4): 579–589. doi : 10.1016/j.bbabio.2011.12.012 . PMID 22236806.
^ Rae T, Goff H (1998). «Гемовая простетическая группа лактопероксидазы. Структурные характеристики гема l и гема l-пептидов». Журнал биологической химии . 273 (43): 27968–27977. doi : 10.1074/jbc.273.43.27968 . PMID 9774411. S2CID 25780396.
^ Purdy M (1983). «Влияние фазы роста и структуры клеточной оболочки на восприимчивость сальмонелл, победивших систему лактопероксидаза-тиоцианат-перекись водорода». Infect. Immun . 39 (3): 1187–95. doi :10.1128/IAI.39.3.1187-1195.1983. PMC 348082. PMID 6341231 .
^ Ohshima H (2003). «Химическая основа канцерогенеза, вызванного воспалением». Arch. Biochem. Biophys . 417 (1): 3–11. doi :10.1016/s0003-9861(03)00283-2. PMID 12921773.
^ Хендерсон Дж. (2003). «Фагоциты производят 5-хлорурацил и 5-бромурацил, два мутагенных продукта миелопероксидазы, в воспалительной ткани человека». J. Biol. Chem . 278 (26): 23522–8. doi : 10.1074/jbc.m303928200 . PMID 12707270. S2CID 19631565.
^ Муршудов Г, Гребенко А, Барынин В, Даутер З, Уилсон К, Вайнштейн Б, Мелик-Адамян В, Браво Дж, Ферран Дж, Феррер Дж., Свитала Дж, Левен ПК, Фита I (1996). «Структура гема d каталаз Penicillium vitale и Escherichia coli» (PDF) . Журнал биологической химии . 271 (15): 8863–8868. дои : 10.1074/jbc.271.15.8863 . PMID 8621527. Архивировано (PDF) из оригинала 24 июля 2018 г.
^ Белевич И (2005). «Оксигенированный комплекс цитохрома bd из Escherichia coli: стабильность и фотолабильность». FEBS Letters . 579 (21): 4567–70. doi :10.1016/j.febslet.2005.07.011. PMID 16087180. S2CID 36465802.
^ Фишер Х, Орт Х (1934). Химия пирролов. Липциг: Ишемия Verlagsgesellschaft.
^ Puustinen A, Wikström M. (1991). "Гемовые группы цитохрома o из Escherichia coli". Proc. Natl. Acad. Sci. USA . 88 (14): 6122–6. Bibcode : 1991PNAS...88.6122P. doi : 10.1073/pnas.88.14.6122 . PMC 52034. PMID 2068092 .
^ Caughey WS, York JL (1962). «Выделение и некоторые свойства зеленого гема цитохромоксидазы из сердечной мышцы говядины». J. Biol. Chem . 237 (7): 2414–6. doi : 10.1016/S0021-9258(19)63456-3 . PMID 13877421.
^ Caughey WS, Smythe GA, O'Keeffe DH, Maskasky JE, Smith ML (1975). «Гем A цитохром c оксидазы. Структура и свойства: сравнение с гемами B, C и S и производными». J. Biol. Chem . 250 (19): 7602–22. doi : 10.1016/S0021-9258(19)40860-0 . PMID 170266.
^ Battersby AR (2000). «Тетрапирролы: пигменты жизни». Natural Product Reports . 17 (6): 507–526. doi :10.1039/B002635M. PMID 11152419.
^ Шридеви К (28 апреля 2018 г.). Повышение регуляции фермента гемового пути ALA-синтазы-1 глютетимидом и 4,6-диоксогептановой кислотой и понижение регуляции глюкозой и гемом: диссертация. EScholarship@UMMS (диссертация). Медицинская школа Массачусетского университета. doi : 10.13028/yyrz-qa79. Архивировано из оригинала 8 августа 2016 г. Получено 28 апреля 2018 г.
^ ab Kumar S, Bandyopadhyay U (июль 2005 г.). «Токсичность свободного гема и ее системы детоксикации у человека». Toxicology Letters . 157 (3): 175–188. doi :10.1016/j.toxlet.2005.03.004. PMID 15917143.
^ Puy H, Gouya L, Deybach JC (март 2010 г.). «Porphyrias». The Lancet . 375 (9718): 924–937. doi :10.1016/S0140-6736(09)61925-5. PMID 20226990. S2CID 208791867.
^ Fraser RZ, Shitut M, Agrawal P, Mendes O, Klapholz S (2018-04-11). «Оценка безопасности соевого леггемоглобина, полученного из Pichia pastoris, предназначенного для использования в качестве катализатора вкуса в растительном мясе». Международный журнал токсикологии . 37 (3): 241–262. doi : 10.1177/1091581818766318. ISSN 1091-5818. PMC 5956568. PMID 29642729 .
^ «Внутри странной науки о поддельном мясе, которое «кровоточит». Wired . 2017-09-20. Архивировано из оригинала 24 марта 2018 года . Получено 28 апреля 2018 года .
^ "Кремниевая долина приобретает вкус к еде". The Economist . 2015-03-05. ISSN 0013-0613 . Получено 2019-04-08 .
^ Maines MD (июль 1988 г.). «Гем-оксигеназа: функция, множественность, регуляторные механизмы и клинические применения». Журнал FASEB . 2 (10): 2557–2568. doi : 10.1096/fasebj.2.10.3290025 . ISSN 0892-6638. PMID 3290025. S2CID 22652094.
^ Ленингер «Принципы биохимии» (5-е изд.). Нью-Йорк: WH Freeman and Company. 2008. С. 876. ISBN978-0-7167-7108-1.
^ Ханафи К (2013). «Окись углерода и мозг: время переосмыслить догму». Curr. Pharm. Des . 19 (15): 2771–5. doi :10.2174/1381612811319150013. PMC 3672861. PMID 23092321 .
^ Абрахам Н., Каппас А. (2008). «Фармакологические и клинические аспекты гем-оксигеназы». Pharmacol. Rev. 60 ( 1): 79–127. doi :10.1124/pr.107.07104. PMID 18323402. S2CID 12792155.
^ Florczyk U, Jozkowicz A, Dulak J (январь–февраль 2008 г.). «Биливердинредуктаза: новые особенности старого фермента и его потенциальное терапевтическое значение». Pharmacological Reports . 60 (1): 38–48. PMC 5536200 . PMID 18276984.
^ King C, Rios G, Green M, Tephly T (2000). «UDP-глюкуронозилтрансферазы». Current Drug Metabolism . 1 (2): 143–161. doi :10.2174/1389200003339171. PMID 11465080.
^ Холл Б., Леви С., Дюфо-Томпсон К., Арп Г., Чжун А., Нджите Г. М., Вайс А., Бракчиа Д., Дженкинс С., Грант М. Р., Абейсингхе С., Ян И., Джермейн М. Д., Ву Ч., Ма Б. (2024-01-03). «BilR — это микробный фермент кишечника, который восстанавливает билирубин до уробилиногена». Nature Microbiology . 9 (1): 173–184. doi : 10.1038/s41564-023-01549-x . ISSN 2058-5276. PMC 10769871. PMID 38172624 .
^ Helmenstine AM. «Химические вещества, ответственные за цвет мочи и фекалий». ThoughtCo . Получено 24.01.2020 .
^ Bunn HF, Jandl JH (сентябрь 1966 г.). «Обмен гема между молекулами гемоглобина». Proc. Natl. Acad. Sci. USA . 56 (3): 974–978. Bibcode : 1966PNAS...56..974B. doi : 10.1073/pnas.56.3.974 . PMC 219955. PMID 5230192.
^ Smith ML, Paul J, Ohlsson PI, Hjortsberg K, Paul KG (февраль 1991 г.). «Расщепление гем-протеина в неденатурирующих условиях». Proc. Natl. Acad. Sci. USA . 88 (3): 882–886. Bibcode :1991PNAS...88..882S. doi : 10.1073/pnas.88.3.882 . PMC 50918 . PMID 1846966.
^ Everse J, Hsia N (1197). «Токсичность нативных и модифицированных гемоглобинов». Free Radical Biology and Medicine . 22 (6): 1075–1099. doi :10.1016/S0891-5849(96)00499-6. PMID 9034247.
^ Кумар С., Бандйопадхай У. (июль 2005 г.). «Токсичность свободного гема и ее системы детоксикации у людей». Toxicology Letters . 157 (3): 175–188. doi :10.1016/j.toxlet.2005.03.004. PMID 15917143.
^ Памплона А, Феррейра А, Балла Дж, Джени В, Балла Г, Эпифанио С, Хора А, Родригес CD, Грегуар IP, Кунья-Родригес М, Португалия С, Соарес MP, Мота ММ (июнь 2007 г.). «Гемоксигеназа-1 и окись углерода подавляют патогенез экспериментальной церебральной малярии». Природная медицина . 13 (6): 703–710. дои : 10.1038/нм1586. PMID 17496899. S2CID 20675040.
^ Ларсен Р., Гоццелино Р., Джени В., Токайи Л., Бозза Ф.А., Джапиасу А.М., Бонапарт Д., Кавальканте М.М., Чора А., Феррейра А., Маргути I, Кардосо С., Сепульведа Н., Смит А., Соареш М.П. (2010). «Центральная роль свободного гема в патогенезе тяжелого сепсиса». Наука трансляционной медицины . 2 (51): 51ра71. doi : 10.1126/scitranslmed.3001118. PMID 20881280. S2CID 423446.
^ Bastide NM, Pierre FH, Corpet DE (февраль 2011 г.). «Гемовое железо из мяса и риск колоректального рака: метаанализ и обзор задействованных механизмов». Cancer Prevention Research (Филадельфия, Пенсильвания) . 4 (2): 177–184. doi :10.1158/1940-6207.CAPR-10-0113. ISSN 1940-6215. PMID 21209396.
^ «Диета, питание, физическая активность и колоректальный рак». wcrf.org. Получено 12 февраля 2022 г.
^ Чанг, Вики С.; Коттерчио, Мишель; Ху, Эдвин (2019). «Потребление железа, уровень железа в организме и риск рака груди: систематический обзор и метаанализ». BMC Cancer . 19 (1): 543. doi : 10.1186/s12885-019-5642-0 . PMC 6555759. PMID 31170936 .{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка )
^ Plewinska M, Thunell S, Holmberg L, Wetmur J, Desnick R (1991). «порфирия с дефицитом дельта-аминолевулинатдегидратазы: идентификация молекулярных поражений у тяжело пораженной гомозиготы». American Journal of Human Genetics . 49 (1): 167–174. PMC 1683193. PMID 2063868 .
^ Аурици С., Лупия Пальмиери Г., Барбьери Л., Макри А., Зорге Ф., Усай Г., Биолкати Г. (февраль 2009 г.). «Четыре новые мутации гена оксидазы копропорфириногена III». Клеточная и молекулярная биология . 55 (1): 8–15. ПМИД 19267996.
^ Bustad HJ, Vorland M, Ronneseth E, Sandberg S, Martinez A, Toska K (8 августа 2013 г.). «Конформационная стабильность и анализ активности двух мутантов гидроксиметилбилансинтазы, K132N и V215E, с различной фенотипической ассоциацией с острой перемежающейся порфирией». Bioscience Reports . 33 (4): 617–626. doi :10.1042/BSR20130045. PMC 3738108 . PMID 23815679.
^ Мартинес ди Монтемурос Ф., Ди Пьерро Э., Патти Э., Тавацци Д., Даниэлли М.Г., Биолкати Г., Рокки Э., Каппеллини, доктор медицины (декабрь 2002 г.). «Молекулярная характеристика порфирий в Италии: схема диагностики». Клеточная и молекулярная биология (Нуази-Ле-Гран, Франция) . 48 (8): 867–876. ISSN 0145-5680. ПМИД 12699245.
^ Badenas C, To Figueras J, Phillips JD, Warby CA, Muñoz C, Herrero C (апрель 2009 г.). «Идентификация и характеристика новых мутаций гена уропорфириногендекарбоксилазы у большого количества пациентов с поздней кожной порфирией и их родственников». Clinical Genetics . 75 (4): 346–353. doi :10.1111/j.1399-0004.2009.01153.x. PMC 3804340 . PMID 19419417.