_(10144905255).jpg/1280px-Anthomyiidae_(female)_(10144905255).jpg)
Мухи — насекомые отряда Diptera , название которого происходит от греческого δι- di- «два» и πτερόν pteron «крыло». Насекомые этого отряда используют для полета только одну пару крыльев, задние крылья превратились в развитые механосенсорные органы, известные как жужжальца , которые действуют как высокоскоростные датчики вращательного движения и позволяют двукрылым выполнять сложные фигуры высшего пилотажа. Diptera — большой отряд, включающий, по оценкам, 1 000 000 видов , включая слепней , [a] долгоножек , журчалок , комаров и других, хотя описано всего около 125 000 видов .
Мухи имеют подвижную голову с парой больших сложных глаз и ротовыми частями, предназначенными для прокалывания и сосания (комары, черные мухи и мухи-грабители) или для лакания и сосания в других группах. Их строение крыльев дает им большую маневренность в полете, а когти и подушечки на ногах позволяют им цепляться за гладкие поверхности. Мухи претерпевают полный метаморфоз ; яйца часто откладываются на личиночном источнике пищи, а личинки, у которых нет настоящих конечностей, развиваются в защищенной среде, часто внутри своего источника пищи. Другие виды являются яйцеживородящими , оппортунистически откладывая вылупившихся или вылупившихся личинок вместо яиц на падаль , навоз, гниющий материал или открытые раны млекопитающих. Куколка представляет собой жесткую капсулу, из которой взрослая особь появляется, когда готова к этому; у мух в основном короткая жизнь во взрослом состоянии.
Двукрылые — один из основных отрядов насекомых, имеющий важное экологическое и человеческое значение. Мухи — важные опылители, уступающие только пчелам и их родственникам -перепончатокрылым . Мухи, возможно, были одними из самых ранних опылителей, ответственных за раннее опыление растений . Плодовые мушки используются в качестве модельных организмов в исследованиях, но менее безвредны: комары являются переносчиками малярии , лихорадки денге , лихорадки Западного Нила , желтой лихорадки , энцефалита и других инфекционных заболеваний ; а комнатные мухи , симбионты людей по всему миру, распространяют пищевые заболевания . Мухи могут быть неприятностями, особенно в некоторых частях мира, где они могут встречаться в больших количествах, жужжа и садясь на кожу или глаза, чтобы укусить или найти жидкости. Более крупные мухи, такие как мухи цеце и мясные личинки, наносят значительный экономический ущерб крупному рогатому скоту. Личинки мясных мух, известные как личинки , и личинки других двукрылых, более известные как личинки , используются в качестве приманки для рыбалки , в качестве пищи для плотоядных животных, а также в медицине для обработки ран .
Двукрылые — это насекомые с полным превращением , которые подвергаются радикальному метаморфозу. Они относятся к Mecopterida , наряду с Mecoptera , Siphonaptera , Lepidoptera и Trichoptera . [3] [4] Наличие одной пары крыльев отличает большинство настоящих мух от других насекомых, в названии которых есть слово «муха». Однако некоторые настоящие мухи, такие как Hippoboscidae (мухи-вши), стали вторично бескрылыми. [5] [6]
Кладограмма представляет текущее консенсусное мнение. [ 7]
.jpg/1280px-Fossil_insect_Diptera,_Brachycera_in_Baltic_amber._Age_50_Mill._years_(the_Lower_Eocene).jpg)
Первые настоящие двукрылые известны со среднего триаса (около 240 миллионов лет назад), и они получили широкое распространение в течение среднего и позднего триаса . [8] Современные цветковые растения не появлялись до мелового периода (около 140 миллионов лет назад), поэтому у первоначальных двукрылых, должно быть, был другой источник питания, отличный от нектара . Основываясь на тяге многих современных групп мух к блестящим каплям, было высказано предположение, что они могли питаться медвяной росой, вырабатываемой сосущими соки клопами , которые были в изобилии в то время, а ротовые части двукрылых хорошо приспособлены к размягчению и лаканию затвердевших остатков. [9] Базальные клады в Diptera включают Deuterophlebiidae и загадочных Nymphomyiidae . [10] Считается, что на основе ископаемых остатков произошло три эпизода эволюционной радиации . Многие новые виды низших Diptera появились в триасе , около 220 миллионов лет назад. Многие низшие Brachycera появились в юрском периоде , около 180 миллионов лет назад. Третья радиация произошла среди Schizophora в начале палеогена , 66 миллионов лет назад. [10]
Филогенетическое положение Diptera было спорным. Монофилия насекомых с полным превращением давно принята, при этом основными отрядами были установлены Lepidoptera, Coleoptera, Hymenoptera и Diptera, и именно отношения между этими группами вызвали трудности. Широко распространено мнение, что Diptera является членом Mecopterida , наряду с Lepidoptera (бабочки и моли), Trichoptera (ручейники), Siphonaptera (блохи), Mecoptera (скорпионницы) и, возможно, Strepsiptera (мухи с закрученными крыльями). Diptera были сгруппированы с Siphonaptera и Mecoptera в Antliophora, но это не было подтверждено молекулярными исследованиями. [11]

Diptera традиционно делились на два подотряда, Nematocera и Brachycera , отличающихся различиями в антеннах. Nematocera идентифицируются по их удлиненным телам и многочлениковым, часто перистым антеннам, как у комаров и долгоножек. Brachycera имеют более округлые тела и гораздо более короткие антенны. [12] [13] Последующие исследования определили Nematocera как немонофилетичных с современными филогениями, помещающими Brachycera в градации групп, ранее помещенных в Nematocera. Построение филогенетического дерева было предметом текущих исследований. Следующая кладограмма основана на проекте FLYTREE. [14] [15]

Мухи часто многочисленны и встречаются почти во всех наземных местообитаниях в мире, за исключением Антарктиды. К ним относятся многие знакомые насекомые, такие как мухи, мясные мухи, комары, мошки, черные мухи, мошки и плодовые мушки. Более 150 000 были формально описаны , и фактическое разнообразие видов намного больше, при этом мухи из многих частей света еще предстоит интенсивно изучить. [16] [17] Подотряд Nematocera включает в себя в основном небольших, тонких насекомых с длинными усиками, таких как комары, мошки, мошки и долгоножки, в то время как Brachycera включает более широких, более крепких мух с короткими усиками. Многие личинки Nematoceran являются водными. [18] По оценкам, в Европе насчитывается около 19 000 видов двукрылых, в Неарктическом регионе — 22 000, в Афротропическом регионе — 20 000, в Восточном регионе — 23 000 и в Австралазийском регионе — 19 000. [19] Хотя большинство видов имеют ограниченное распространение, некоторые, например, муха домашняя ( Musca domestica ), являются космополитами. [20] Gauromydas heros ( Asiloidea ), длиной до 7 см (2,8 дюйма), обычно считается самой большой мухой в мире, [21] а самой маленькой является Euryplatea nanaknihali , которая при длине 0,4 мм (0,016 дюйма) меньше крупинки соли. [22]
Brachycera экологически очень разнообразны, многие из них являются хищниками на личиночной стадии, а некоторые являются паразитами. Паразитирующими животными являются моллюски , мокрицы , многоножки , насекомые, млекопитающие [ 19] и земноводные [23] . Мухи являются второй по величине группой опылителей после перепончатокрылых (пчелы, осы и родственники). Во влажных и холодных условиях мухи значительно более важны как опылители. По сравнению с пчелами, им нужно меньше пищи, поскольку им не нужно обеспечивать свое потомство. Многие цветы, которые несут мало нектара, и те, которые развили опыление ловушками, зависят от мух. [24] Считается, что некоторые из самых ранних опылителей растений, возможно, были мухами [25] .
Наибольшее разнообразие галлообразующих насекомых встречается среди мух, в основном в семействе Cecidomyiidae (галлицы). [26] Многие мухи (главным образом в семействе Agromyzidae) откладывают яйца в мезофильную ткань листьев, а личинки питаются между поверхностями, образуя волдыри и мины. [27] Некоторые семейства являются микофагами или грибоядными. К ним относятся пещерные Mycetophilidae (грибные комарики), личинки которых являются единственными двукрылыми с биолюминесценцией. Sciaridae также являются грибоядными. Некоторые растения опыляются грибоядными мухами, которые посещают зараженные грибком мужские цветки. [28]
Личинки Megaselia scalaris (Phoridae) почти всеядны и потребляют такие вещества, как краска и крем для обуви. [29] Муха Exorista mella (Walker) считается универсалом и паразитоидом различных хозяев. [30] Личинки береговых мух (Ephydridae) и некоторые Chironomidae выживают в экстремальных условиях, включая ледники ( Diamesa sp., Chironomidae [31] ), горячие источники, гейзеры, соленые бассейны, серные бассейны , септики и даже сырую нефть ( Helaeomyia petrolei [31] ). [19] Взрослые журчалки (Syrphidae) хорошо известны своей мимикрией , а личинки ведут разнообразный образ жизни, в том числе питаясь инквилинами внутри гнезд общественных насекомых. [32] Некоторые короткоусые ракообразные являются сельскохозяйственными вредителями, некоторые кусают животных и людей и сосут их кровь, а некоторые переносят болезни. [19]
Мухи приспособлены к воздушному движению и обычно имеют короткие и обтекаемые тела. Первая тагма мухи, голова, несет глаза, усики и ротовые части (верхняя губа, нижняя губа, нижняя челюсть и верхняя челюсть составляют ротовые части). Вторая тагма, грудь , несет крылья и содержит летательные мышцы на втором сегменте, который значительно увеличен; первый и третий сегменты были редуцированы до воротникообразных структур, а третий сегмент несет жужжальца , которые помогают насекомому балансировать во время полета. Третья тагма — брюшко , состоящее из 11 сегментов, некоторые из которых могут быть слиты, а 3 самых задних сегмента модифицированы для размножения. [33] [34] Некоторые двукрылые являются имитаторами и могут быть отличимы от своих моделей только при очень тщательном осмотре. Примером этого является Spilomyia longicornis , которая является мухой, но имитирует осу-веспиду .
.jpg/1280px-Tabanus_atratus,_U,_Face,_MD_2013-08-21-16.06.31_ZS_PMax_(9599360121).jpg)
_(25234095874).jpg/1280px-Tephrochlamys_rufiventris_(female_head)_(25234095874).jpg)
Мухи имеют подвижную голову с парой больших сложных глаз по бокам головы, а у большинства видов — три маленьких глазка наверху. Сложные глаза могут быть близко друг к другу или широко разделены, а в некоторых случаях разделены на дорсальную и вентральную области, возможно, для помощи в роевом поведении. Усики хорошо развиты, но изменчивы, будучи нитевидными, перистыми или гребневидными в разных семействах. Ротовые части приспособлены для прокалывания и сосания, как у черных мух, комаров и мух-грабителей, а также для лакания и сосания, как у многих других групп. [34] Самки слепней используют ножевидные мандибулы и максиллы, чтобы сделать крестообразный надрез на коже хозяина, а затем лакать вытекающую кровь. Кишечник включает большие дивертикулы , позволяющие насекомому хранить небольшое количество жидкости после еды. [35]
Для визуального контроля курса поле оптического потока мух анализируется набором нейронов, чувствительных к движению. [36] Предполагается, что подмножество этих нейронов участвует в использовании оптического потока для оценки параметров собственного движения, таких как рыскание, крен и боковое перемещение. [37] Предполагается, что другие нейроны участвуют в анализе содержания самой визуальной сцены, например, в отделении фигур от земли с помощью параллакса движения. [38] [39] Нейрон H1 отвечает за обнаружение горизонтального движения по всему полю зрения мухи, позволяя мухе генерировать и направлять стабилизирующие двигательные коррекции в полете относительно рыскания. [40] Глазки участвуют в обнаружении изменений интенсивности света, позволяя мухе быстро реагировать на приближение объекта. [41]
Как и другие насекомые, мухи имеют хеморецепторы , которые определяют запах и вкус, и механорецепторы , которые реагируют на прикосновение. Третьи сегменты антенн и верхнечелюстные щупики несут основные обонятельные рецепторы, в то время как вкусовые рецепторы находятся в губе, глотке, ногах, краях крыльев и женских гениталиях, [42] позволяя мухам пробовать пищу, ходя по ней. Вкусовые рецепторы у самок на кончике брюшка получают информацию о пригодности места для откладки яиц. [41] Мухи, питающиеся кровью, имеют специальные сенсорные структуры, которые могут обнаруживать инфракрасное излучение и использовать его для наведения на своих хозяев. Многие кровососущие мухи могут обнаруживать повышенную концентрацию углекислого газа , которая возникает вблизи крупных животных. [43] Некоторые тахиновые мухи (Ormiinae), которые являются паразитоидами кустарниковых сверчков , имеют звуковые рецепторы, которые помогают им находить своих поющих хозяев. [44]

У двукрылых одна пара передних крыльев на среднегруди и пара жужжальца , или редуцированных задних крыльев, на заднегруди . Дальнейшая адаптация к полету — это уменьшение количества нервных ганглиев и концентрация нервной ткани в грудной клетке, особенность, которая наиболее выражена в высокопроизводном инфраотряде Muscomorpha. [35] Некоторые мухи, такие как эктопаразитические Nycteribiidae и Streblidae, являются исключением, поскольку утратили крылья и стали нелетающими. Единственный другой отряд насекомых, несущий одну пару настоящих функциональных крыльев, в дополнение к любой форме жужжальца, — это Strepsiptera . В отличие от мух, Strepsiptera несут свои жужжальца на среднегруди, а свои маховые крылья — на заднегруди. [45] Каждая из шести ног мухи имеет типичную для насекомых структуру: тазик, вертел, бедренная кость, большая берцовая кость и цевка, причем цевка в большинстве случаев подразделяется на пять тарсомеров . [34] На конце конечности находится пара коготков, а между ними находятся подушкообразные структуры, известные как пульвиллы , которые обеспечивают сцепление. [46]
Брюшко показывает значительную изменчивость среди членов отряда. Оно состоит из одиннадцати сегментов в примитивных группах и десяти сегментов в более производных группах, десятый и одиннадцатый сегменты слились. [47] Последние два или три сегмента приспособлены для размножения. Каждый сегмент состоит из спинного и вентрального склерита , соединенных эластичной мембраной. У некоторых самок склериты свернуты в гибкий телескопический яйцеклад . [34]

Мухи способны к большой маневренности во время полета благодаря наличию жужжальца. Они действуют как гироскопические органы и быстро колеблются в такт с крыльями; они действуют как система баланса и наведения, обеспечивая быструю обратную связь с мышцами, управляющими крыльями, и мухи, лишенные жужжальца, не способны летать. Крылья и жужжальца движутся синхронно, но амплитуда каждого взмаха крыла независима, что позволяет мухе поворачиваться вбок. [48] Крылья мухи прикреплены к двум видам мышц: тем, которые используются для ее питания, и другому набору, используемому для точного управления. [49]
Мухи, как правило, летают по прямой линии, а затем быстро меняют направление, прежде чем продолжить полет по другой прямой траектории. Изменения направления называются саккадами и обычно включают угол в 90°, достигаемый за 50 миллисекунд. Они инициируются визуальными стимулами, когда муха наблюдает за объектом, затем нервы активируют рулевые мышцы в грудной клетке, которые вызывают небольшое изменение в взмахе крыльев, что создает достаточный крутящий момент для поворота. Обнаружив это в течение четырех или пяти взмахов крыльев, жужжальца запускают встречный поворот, и муха направляется в новом направлении. [50]
У мух быстрые рефлексы, которые помогают им убежать от хищников, но их постоянная скорость полета низкая. Мухи -долихоподы рода Condylostylus реагируют на вспышки фотокамер менее чем за 5 миллисекунд, взлетая. [51] В прошлом олений овод Cephenemyia считался одним из самых быстрых насекомых на основе визуальной оценки, сделанной Чарльзом Таунсендом в 1927 году. [52] Это утверждение о скорости от 600 до 800 миль в час регулярно повторялось, пока Ирвинг Ленгмюр не доказал, что оно физически невозможно, а также неверно. Ленгмюр предположил расчетную скорость в 25 миль в час. [53] [54] [55]
Хотя большинство мух живут и летают близко к земле, известно, что некоторые из них летают на высоте, а некоторые, такие как Oscinella (Chloropidae), переносятся ветром на высоте до 2000 футов и на большие расстояния. [56] Известно, что некоторые журчащие мухи, такие как Metasyrphus corollae, совершают длительные перелеты в ответ на всплески популяции тли. [57]
Самцы видов мух, таких как Cuterebra , многие журчалки, [58] пчелиные мухи (Bombyliidae) [59] и плодовые мушки (Tephritidae) [60] сохраняют территории, в пределах которых они занимаются воздушным преследованием, чтобы отогнать вторгшихся самцов и другие виды. [61] Хотя эти территории могут удерживаться отдельными самцами, некоторые виды, такие как A. freeborni , [62] образуют токовища со множеством самцов, собирающихся в группы. [60] Некоторые мухи сохраняют воздушное пространство, а другие образуют плотные стаи, которые сохраняют неподвижное местоположение относительно ориентиров. Многие мухи спариваются в полете во время роения. [63]
.jpg/1280px-Anthomyiidae_sp._1_(aka).jpg)
Двукрылые насекомые проходят полный метаморфоз, в ходе которого они проходят четыре четко выраженные стадии развития: яйцо, личинка, куколка и взрослая особь.
У многих мух личиночная стадия длинная, а у взрослых особей может быть короткая жизнь. Большинство личинок двукрылых развиваются в защищенной среде; многие из них водные, а другие встречаются во влажных местах, таких как падаль, фрукты, овощи, грибы и, в случае паразитических видов, внутри своих хозяев. Они, как правило, имеют тонкую кутикулу и высыхают при контакте с воздухом. За исключением Brachycera , большинство личинок двукрылых имеют склеротизированные головные капсулы, которые могут быть редуцированы до остатков ротовых крючков; однако Brachycera имеют мягкие, желатинизированные головные капсулы, в которых склериты редуцированы или отсутствуют. Многие из этих личинок втягивают голову в грудную клетку. [34] [64] Дыхальца личинки и куколки не имеют внутреннего механического запирающего устройства. [65]

Некоторые другие анатомические различия существуют между личинками Nematocera и Brachycera . Особенно у Brachycera, между грудью и брюшком наблюдается небольшая демаркация, хотя демаркация может быть видна у многих Nematocera, таких как комары; у Brachycera голова личинки нечетко отличается от остальной части тела, и присутствует мало, если вообще присутствует, склеритов. Неформально такие личинки brachycera называются личинками, [66] но этот термин не является техническим и часто применяется безразлично к личинкам мух или личинкам насекомых в целом. Глаза и усики личинок brachycera редуцированы или отсутствуют, а на брюшке также отсутствуют придатки, такие как церки . Это отсутствие признаков является адаптацией к пище, такой как падаль, разлагающийся детрит или ткани хозяина, окружающие эндопаразитов . [35] Личинки Nematoceran обычно имеют хорошо развитые глаза и усики, тогда как у личинок Brachyceran они редуцированы или модифицированы. [67]
Личинки двукрылых не имеют сочлененных, «настоящих ног», [64] но некоторые личинки двукрылых, такие как виды Simuliidae , Tabanidae и Vermileonidae , имеют ложные ноги, приспособленные для удержания субстрата в текущей воде, тканей хозяина или добычи. [68] Большинство двукрылых являются яйцекладущими и откладывают партии яиц, но некоторые виды являются яйцеживородящими , когда личинки начинают развиваться внутри яиц до того, как они вылупятся, или живородящими, когда личинки вылупляются и созревают в теле матери, прежде чем будут отложены наружу. Они встречаются особенно в группах, в которых личинки зависят от источников пищи, которые являются недолговечными или доступны в течение коротких периодов времени. [69] Это широко распространено в некоторых семействах, таких как Sarcophagidae. У Hylemya strigosa (Anthomyiidae) личинка линяет до второй стадии перед вылуплением, а у Termitoxenia (Phoridae) самки имеют инкубационные сумки, а полностью развитая личинка третьей стадии откладывается взрослой особью, и она почти сразу окукливается без свободно питающейся личиночной стадии. Муха цеце (а также другие Glossinidae, Hippoboscidae, Nycteribidae и Streblidae) демонстрирует аденотрофное живорождение ; одно оплодотворенное яйцо остается в яйцеводе, а развивающаяся личинка питается железистыми выделениями. Когда самка полностью вырастает, она находит место с мягкой почвой, и личинка пробирается из яйцевода, закапывается и окукливается. Некоторые мухи, такие как Lundstroemia parthenogenetica (Chironomidae), размножаются путем телитокического партеногенеза , а некоторые галлицы имеют личинки, которые могут производить яйца ( педогенез ). [70] [71]
Куколки принимают различные формы. В некоторых группах, особенно Nematocera, куколка является промежуточной между личиночной и взрослой формой; эти куколки описываются как «обтектные», имея будущие придатки, видимые как структуры, которые прилипают к телу куколки. Внешняя поверхность куколки может быть кожистой и иметь шипы, дыхательные особенности или локомоторные лопасти. В других группах, описываемых как «коарктированные», придатки не видны. В них внешняя поверхность представляет собой пупарий , образованный из последней личиночной кожи, а фактическая куколка скрыта внутри. Когда взрослое насекомое готово выйти из этой жесткой, устойчивой к высыханию капсулы, оно надувает на голове похожую на шар структуру и проталкивается наружу. [34]
Стадия взрослой особи обычно короткая, ее функция заключается только в спаривании и откладывании яиц. Гениталии самцов мух повернуты в разной степени по сравнению с положением, обнаруженным у других насекомых. [72] У некоторых мух это временное вращение во время спаривания, но у других это постоянное скручивание органов, которое происходит во время стадии куколки. Это скручивание может привести к тому, что анус окажется ниже гениталий, или, в случае скручивания на 360°, к тому, что семенной проток будет обернут вокруг кишечника, а внешние органы окажутся в своем обычном положении. Когда мухи спариваются, самец сначала летит сверху самки, лицом в том же направлении, но затем разворачивается лицом в противоположном направлении. Это заставляет самца лечь на спину, чтобы его гениталии оставались связанными с гениталиями самки, или скручивание гениталий самца позволяет самцу спариваться, оставаясь в вертикальном положении. Это приводит к тому, что мухи имеют больше репродуктивных способностей, чем большинство насекомых, и гораздо быстрее. Мухи встречаются в больших популяциях из-за их способности эффективно и быстро спариваться во время брачного сезона. [35] Более примитивные группы спариваются в воздухе во время роения, но большинство более продвинутых видов с 360° торсионом спариваются на субстрате. [73]

Как вездесущие насекомые, двукрылые играют важную роль на различных трофических уровнях как потребители и как добыча. В некоторых группах личинки завершают свое развитие без питания, а в других взрослые особи не питаются. Личинки могут быть травоядными, падальщиками, редуцентами, хищниками или паразитами, причем потребление разлагающейся органики является одним из наиболее распространенных видов пищевого поведения. Фрукты или детрит потребляются вместе с сопутствующими микроорганизмами, ситообразный фильтр в глотке используется для концентрации частиц, в то время как плотоядные личинки имеют ротовые крючки, которые помогают измельчать пищу. Личинки некоторых групп питаются живыми тканями растений и грибов или в них, и некоторые из них являются серьезными вредителями сельскохозяйственных культур. Некоторые водные личинки потребляют пленки водорослей, которые образуются под водой на камнях и растениях. Многие из паразитоидных личинок растут внутри и в конечном итоге убивают других членистоногих, в то время как паразитические личинки могут нападать на позвоночных хозяев. [34]
В то время как многие личинки двукрылых обитают в воде или в закрытых наземных местах, большинство взрослых особей живут над землей и способны летать. В основном они питаются нектаром или растительными или животными экссудатами, такими как медвяная роса, для чего приспособлены их лакающие ротовые органы. У некоторых мух есть функциональные мандибулы, которые могут использоваться для укусов. У мух, питающихся кровью позвоночных, есть острые стилеты, которые прокалывают кожу, а у некоторых видов есть антикоагулянтная слюна, которая отрыгивается перед поглощением вытекающей крови; в этом процессе могут передаваться определенные заболевания. Оводы (Oestridae) эволюционировали, чтобы паразитировать на млекопитающих. Многие виды завершают свой жизненный цикл внутри тел своих хозяев. [74] Личинки нескольких групп мух (Agromyzidae, Anthomyiidae, Cecidomyiidae) способны вызывать образование галлов на растениях. Некоторые личинки двукрылых являются листовыми минерами. Личинки многих семейств короткоусых являются хищными. Во многих группах двукрылых роение является особенностью взрослой жизни, когда тучи насекомых собираются в определенных местах; эти насекомые в основном самцы, и рой может служить цели сделать их местоположение более заметным для самок. [34]
У большинства взрослых двукрылых ротовые части модифицированы для впитывания жидкости. Взрослые особи многих видов мух (например, Anthomyia sp., Steganopsis melanogaster ), которые питаются жидкой пищей, будут отрыгивать жидкость, что называется «пузырением», что, как полагают, помогает насекомым испарять воду и концентрировать пищу [75] или, возможно, охлаждаться путем испарения. [76] Некоторые взрослые двукрылые известны клептопаразитизмом , например, представители Sarcophagidae. Мильтограммины известны как «мухи-спутники» за их привычку следовать за осами и красть их ужаленную добычу или откладывать в них яйца. Известно, что фориды, милихиды и род Bengalia крадут еду, которую несут муравьи. [77] Взрослые особи Ephydra hians добывают корм под водой и имеют специальные гидрофобные волоски, которые улавливают пузырь воздуха, позволяющий им дышать под водой. [78]

Мухи поедаются другими животными на всех стадиях их развития. Яйца и личинки паразитируют на других насекомых и поедаются многими существами, некоторые из которых специализируются на питании мухами, но большинство из них потребляют их как часть смешанной диеты. Птицы, летучие мыши, лягушки, ящерицы, стрекозы и пауки являются хищниками мух. [79] Многие мухи развили миметическое сходство , которое помогает им защищаться. Бейтсовская мимикрия широко распространена у многих журчалок, напоминающих пчел и ос, [80] [81] муравьев [82] и некоторых видов плодовых мух-тефритид, напоминающих пауков. [83] Некоторые виды журчалок являются мирмекофилами — их детеныши живут и растут в гнездах муравьев. Они защищены от муравьев, имитируя химические запахи, издаваемые членами колонии муравьев. [84] Пчелиные мухи-бомбилииды, такие как Bombylius major, имеют короткое тело, круглую форму, покрыты пушистым мехом и отчетливо напоминают пчел, поскольку они посещают цветы в поисках нектара, и, вероятно, также являются бейтсовскими имитаторами пчел. [85]
Напротив, у Drosophila subobscura , вида мух из рода Drosophila , отсутствует категория гемоцитов, которые присутствуют у других изученных видов Drosophila , что приводит к неспособности защищаться от атак паразитов, что является формой врожденного иммунодефицита. [86]

Мухи играют множество символических ролей в разных культурах. Они включают как положительные, так и отрицательные роли в религии. В традиционной религии навахо Большая Муха является важным духовным существом. [87] [88] [89] В христианской демонологии Вельзевул — демоническая муха , «Повелитель мух» и бог филистимлян . [ 90] [91] [92]
Мухи появились в литературе со времен Древнего Шумера . [93] В шумерской поэме муха помогает богине Инанне , когда ее мужа Думузида преследуют демоны галла . [93] В месопотамских версиях мифа о потопе мертвые тела, плавающие в водах, сравниваются с мухами. [93] Позже говорится, что боги роятся «как мухи» вокруг подношения героя Утнапиштима . [93] Мухи появляются на старых вавилонских печатях как символы Нергала , бога смерти. [93] В древней Месопотамии часто носили бусины из лазурита в форме мух , наряду с другими видами украшений в виде мух. [93]
В Древнем Египте мухи появляются в амулетах и как военная награда за храбрость и упорство, из-за того, что они всегда возвращаются, если их прихлопнуть. Считается, что мухи также могли быть связаны с отлетающим духом мертвых, так как их часто можно встретить рядом с мертвыми телами. В современном Египте в некоторых областях сохраняется похожее поверье: не прихлопывать блестящих зеленых мух, так как они могут нести душу недавно умершего человека. [94]
В малоизвестном греческом мифе очень болтливая и говорливая дева по имени Мия (что означает «муха») разгневала богиню луны Селену , пытаясь соблазнить ее возлюбленного, спящего Эндимиона , и была превращена разгневанной богиней в муху, которая теперь всегда лишает людей сна в память о своей прошлой жизни. [95] [96] В «Прикованном Прометее » , который приписывается афинскому трагику Эсхилу , овод , посланный женой Зевса Герой , преследует и мучает его любовницу Ио , которая была превращена в корову и за которой постоянно следят сто глаз пастуха Аргуса : [97] [98] «Ио: Ах! Ха! Опять укол, укол жала овода! О земля, земля, скройся, полая форма — Аргус — эта злая вещь — стоглазая». [98] Уильям Шекспир , вдохновленный Эсхилом, создал Тома из Бедлама в «Короле Лире» , «которого нечистый дьявол провел сквозь огонь и пламя, через брод и водоворот, через болото и трясину», сведенного с ума постоянным преследованием. [98] В «Антонии и Клеопатре» Шекспир аналогичным образом сравнивает поспешный отъезд Клеопатры с поля битвы при Акциуме с отъездом коровы, преследуемой оводом. [99] Совсем недавно, в 1962 году, биолог Винсент Детье написал книгу « Знать муху» , познакомив широкого читателя с поведением и физиологией мухи. [100]
Musca describea («нарисованная муха» на латыни) — изображение мухи как незаметного элемента различных картин. Эта особенность была широко распространена в картинах XV и XVI веков, и ее присутствие может быть объяснено разными причинами. [101]
Мухи появляются в популярной культуре в таких концепциях, как документальное кино «мухи на стене» в кино- и телепроизводстве . Метафорическое название предполагает, что события рассматриваются откровенно , как их могла бы видеть муха. [102] Мухи вдохновили на создание миниатюрных летающих роботов. [103] Фильм Стивена Спилберга 1993 года « Парк Юрского периода» основывался на идее о том, что ДНК может сохраняться в содержимом желудка кровососущей мухи, окаменевшей в янтаре , хотя этот механизм был отвергнут учеными. [104]
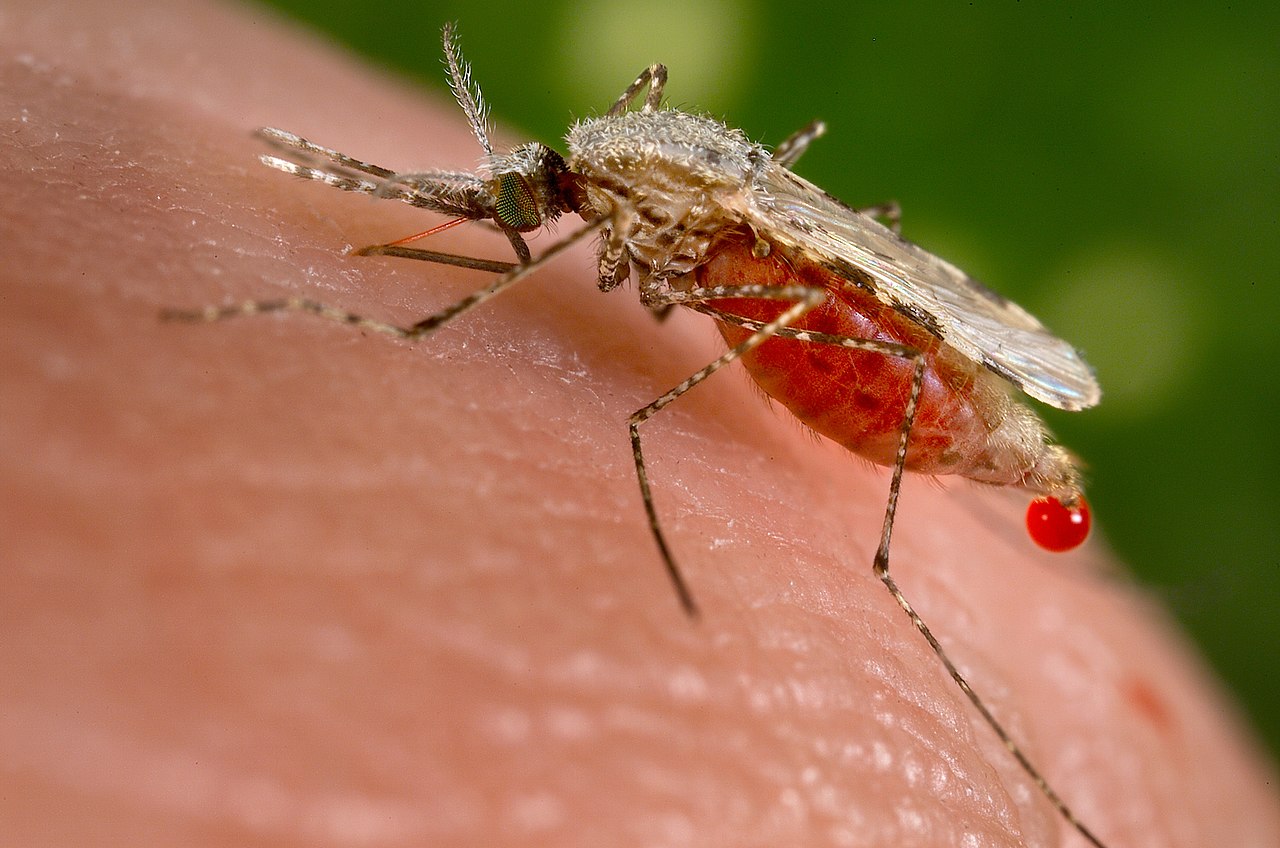
Двукрылые — важная группа насекомых, оказывающая значительное влияние на окружающую среду. Некоторые минирующие мухи (Agromyzidae), плодовые мухи (Tephritidae и Drosophilidae) и галлицы (Cecidomyiidae) являются вредителями сельскохозяйственных культур; другие, такие как мухи цеце , мясные личинки и оводы (Oestridae), нападают на скот, вызывая раны, распространяя болезни и нанося значительный экономический ущерб. См. статью: Паразитические мухи домашних животных . Некоторые из них могут даже вызывать миазы у людей. Другие, такие как комары (Culicidae), мошки (Simuliidae) и дренажные мухи (Psychodidae), влияют на здоровье человека, выступая в качестве переносчиков основных тропических заболеваний. Среди них комары Anopheles переносят малярию , филяриатоз и арбовирусы ; комары Aedes aegypti переносят лихорадку денге и вирус Зика ; Мошки переносят речную слепоту ; песчаные мухи переносят лейшманиоз . Другие двукрылые доставляют неудобства человеку, особенно когда присутствуют в больших количествах; к ним относятся комнатные мухи, которые загрязняют пищу и распространяют пищевые заболевания; мокрецы и москиты (Ceratopogonidae), а также комнатные мухи и мухи-охвостни (Muscidae). [34] В тропических регионах глазные мухи ( Chloropidae ), которые посещают глаза в поисках жидкости, могут быть неприятностью в некоторые сезоны. [105]
Многие двукрылые выполняют функции, полезные для человека. Мухи комнатные, мясные и грибные комарики (Mycetophilidae) являются падальщиками и способствуют разложению. Мухи-грабители (Asilidae), тахины (Tachinidae), мухи-кинжалы и мухи-воздушные шары (Empididae) являются хищниками и паразитоидами других насекомых, помогая контролировать множество вредителей. Многие двукрылые, такие как пчелиные мухи (Bombyliidae) и журчалки (Syrphidae), являются опылителями сельскохозяйственных культур. [34]

Drosophila melanogaster , плодовая мушка, долгое время использовалась в качестве модельного организма в исследованиях из-за легкости, с которой ее можно разводить и выращивать в лабораторных условиях, ее небольшого генома и того факта, что многие из ее генов имеют аналоги у высших эукариот . На основе этого вида было проведено большое количество генетических исследований; они оказали глубокое влияние на изучение экспрессии генов , механизмов регуляции генов и мутаций . Другие исследования изучали физиологию , микробный патогенез и развитие среди других тем исследований. [106] Исследования двукрылых Вилли Хеннига помогли в разработке кладистики , методов, которые он применял к морфологическим признакам, но теперь адаптированы для использования с молекулярными последовательностями в филогенетике. [107]
Личинки , обнаруженные на трупах, полезны для судебных энтомологов . Виды личинок можно идентифицировать по их анатомическим особенностям и путем сопоставления их ДНК . Личинки разных видов мух посещают трупы и туши в довольно четко определенное время после смерти жертвы, как и их хищники, такие как жуки семейства Histeridae . Таким образом, присутствие или отсутствие определенных видов дает доказательства времени с момента смерти, а иногда и другие детали, такие как место смерти, когда виды ограничены определенными местообитаниями, такими как лесная местность . [108]

Некоторые виды личинок, такие как личинки мясной мухи (gentles) и личинки синей мухи ( casters ), разводятся в коммерческих целях; они продаются в качестве приманки при ловле рыбы и в качестве пищи для плотоядных животных (содержащихся в качестве домашних животных, в зоопарках или для исследований), таких как некоторые млекопитающие , [109] рыбы , рептилии и птицы . Было высказано предположение, что личинки мух могут использоваться в больших масштабах в качестве пищи для выращиваемых на фермах кур, свиней и рыб. Однако потребители выступают против включения насекомых в свою пищу, а использование насекомых в кормах для животных остается незаконным в таких регионах, как Европейский Союз . [110] [111]
Личинки мух могут использоваться в качестве биомедицинского инструмента для ухода за ранами и их лечения. Терапия личинками (MDT) — это использование личинок мясных мух для удаления омертвевших тканей из ран, чаще всего ампутаций. Исторически это использовалось на протяжении столетий, как преднамеренно, так и непреднамеренно, на полях сражений и в ранних больничных условиях. [112] Удаление омертвевших тканей способствует росту клеток и здоровому заживлению ран. Личинки также обладают биохимическими свойствами, такими как антибактериальная активность, обнаруженная в их выделениях во время питания. [113] Эти лекарственные личинки являются безопасным и эффективным средством лечения хронических ран. [114]
Сардинский сыр casu marzu подвергается воздействию мух, известных как сырные прыгуны, таких как Piophila casei , члены семейства Piophilidae . [115] Пищеварительная деятельность личинок мух смягчает сыр и изменяет аромат в процессе созревания. Одно время власти Европейского Союза запретили продажу сыра, и его становилось трудно найти, [ 116] но запрет был снят на том основании, что сыр является традиционным местным продуктом, изготовленным традиционными методами. [117]
Мухи опасны для здоровья и их привлекает запах туалетов. Журнал New Scientist предложил ловушку для этих мух. К туалету была прикреплена труба, действующая как дымоход, которая пропускала немного света, чтобы привлечь этих мух к концу этой трубы, где марлевое покрытие не позволяло им выбраться наружу, так что они оказывались в ловушке и погибали. Туалеты, как правило, темные внутри, особенно если дверь закрыта.
{{cite book}}: CS1 maint: multiple names: authors list (link)Почти каждый элемент во вселенной может быть персонализирован таким образом, и даже самые незначительные из них, такие как крошечный бурундук и те маленькие насекомые, помощники и наставники божества и человека в мифах, Большая Муха (
Dǫ'soh
) и Девочка-Кукурузник ('
Anilt'ánii 'At'ééd
) (Wyman and Bailey 1964:29–30, 51, 137–144), так же необходимы для гармоничного равновесия вселенной, как и великое Солнце.
Общий
Анатомия
Описатели