Губки (также известные как морские губки ), члены типа Porifera ( / p ə ˈ r ɪ f ər ə / ; что означает «носитель пор»), являются базальной кладой животных как сестра дипобластов . [2] [3] [4] [5] [6] Это многоклеточные организмы, тела которых полны пор и каналов, позволяющих воде циркулировать через них, состоящих из желеобразного мезоила , зажатого между двумя тонкими слоями клеток .
Губки имеют неспециализированные клетки, которые могут трансформироваться в другие типы и при этом часто мигрируют между основными слоями клеток и мезохилом. У губок нет сложной нервной , [7] пищеварительной или кровеносной систем, как у человека. Вместо этого большинство полагаются на поддержание постоянного потока воды через свое тело для получения пищи и кислорода, а также для удаления отходов. Губки, которые считаются одними из самых ранних животных, живущих сегодня, возможно, были первыми, кто ответвился на эволюционном древе от последнего общего предка всех животных, что сделало их сестринской группой всех других животных. [2]
Термин «губка» происходит от древнегреческого слова σπόγγος ( spóngos «губка»). [8]

Губки похожи на других животных тем, что они многоклеточные , гетеротрофные , лишены клеточных стенок и производят сперматозоиды . В отличие от других животных, у них отсутствуют настоящие ткани [9] и органы . [10] Некоторые из них радиально симметричны, но большинство асимметричны. Формы их тел приспособлены для максимальной эффективности потока воды через центральную полость, где вода откладывает питательные вещества, а затем выходит через отверстие, называемое оскулумом . Многие губки имеют внутренние скелеты спикул (скелетоподобные фрагменты карбоната кальция или диоксида кремния ) и/или спонгина (модифицированный тип белка коллагена). [9] Все взрослые губки являются сидячими водными животными, что означает, что они прикрепляются к подводной поверхности и остаются фиксированными на месте (т. е. не перемещаются), в то время как на личиночной стадии жизни они подвижны . Хотя существуют пресноводные виды, подавляющее большинство из них являются морскими (солеными) видами, среда обитания которых варьируется от приливных зон до глубин, превышающих 8800 м (5,5 миль).
Хотя большинство из примерно 5 000–10 000 известных видов губок питаются бактериями и другой микроскопической пищей в воде, некоторые из них являются хозяевами фотосинтезирующих микроорганизмов в качестве эндосимбионтов , и эти союзы часто производят больше пищи и кислорода, чем потребляют. Некоторые виды губок, обитающие в условиях скудной пищи, превратились в хищников , питающихся в основном мелкими ракообразными . [11]
Губки размножаются как бесполым, так и половым путем. Большинство видов, использующих половое размножение, выпускают сперматозоиды в воду для оплодотворения яйцеклеток , которые у некоторых видов высвобождаются, а у других сохраняются «матерью». Из оплодотворенных яиц развиваются личинки , которые уплывают в поисках места для поселения. [12] Известно, что губки регенерируют из отколовшихся фрагментов, хотя это работает только в том случае, если фрагменты содержат клетки нужного типа. Некоторые виды размножаются почкованием. Когда условия окружающей среды становятся менее благоприятными для губок, например, при понижении температуры, многие пресноводные виды и некоторые морские виды производят геммулы , «стручки выживания» из неспециализированных клеток, которые остаются в состоянии покоя до тех пор, пока условия не улучшатся; затем они либо образуют совершенно новые губки, либо повторно заселяют скелеты своих родителей. [13]
У большинства губок внутренняя студенистая матрица, называемая мезогилом, функционирует как эндоскелет , и это единственный скелет мягких губок, который покрывает такие твердые поверхности, как камни. Чаще всего мезохил укрепляется минеральными спикулами , спонгиновыми волокнами или тем и другим. Демогубки используют спонгин; многие виды имеют спикулы из кремнезема , тогда как некоторые виды имеют экзоскелет из карбоната кальция . Демогубки составляют около 90% всех известных видов губок, включая все пресноводные, и имеют широчайший ареал обитания. Известковые губки , имеющие спикулы из карбоната кальция, а у некоторых видов и экзоскелеты из карбоната кальция, обитают в относительно мелких морских водах, где добыча карбоната кальция проще всего. [14] Хрупкие стеклянные губки с « строительными лесами » из спикул кремнезема обитают только в полярных регионах и на глубинах океана, где хищники редки. Окаменелости всех этих типов были найдены в породах, датируемых 580 миллионами лет назад . Кроме того, археоциатиды , чьи окаменелости встречаются в горных породах от 530 до 490 миллионов лет назад , теперь считаются разновидностью губок.

Одноклеточные хоанофлагелляты напоминают клетки хоаноцитов губок, которые используются для управления их системами потока воды и захвата большей части пищи . Это, наряду с филогенетическими исследованиями рибосомальных молекул, было использовано в качестве морфологического доказательства, позволяющего предположить, что губки являются сестринской группой по отношению к остальным животным. [16]
Несколько видов демоспонги, которые имеют полностью мягкий волокнистый скелет без твердых элементов, использовались людьми на протяжении тысячелетий для различных целей, в том числе в качестве прокладки и инструментов для чистки. Однако к 1950-м годам они были настолько переловлены , что отрасль почти рухнула, и большинство подобных губке материалов теперь являются синтетическими. Губки и их микроскопические эндосимбионты сейчас исследуются как возможные источники лекарств для лечения широкого спектра заболеваний. Было замечено, что дельфины используют губки в качестве инструментов во время добывания пищи . [17]
Губки составляют тип Porifera и определяются как сидячие многоклеточные животные (многоклеточные неподвижные животные), у которых есть входные и выходные отверстия для воды, соединенные камерами, выстланными хоаноцитами , клетками с хлыстовидными жгутиками. [18] Однако некоторые плотоядные губки потеряли эти системы потока воды и хоаноциты. [19] [20] Все известные живые губки могут переделывать свое тело, поскольку большинство типов их клеток могут перемещаться внутри тела, а некоторые могут переходить из одного типа в другой. [20] [21]
Даже если несколько губок способны производить слизь, которая действует как микробный барьер у всех других животных, ни одна губка, способная выделять функциональный слой слизи, не зарегистрирована. Без такого слоя слизи их живая ткань покрыта слоем микробных симбионтов, которые могут составлять до 40–50% влажной массы губок. Эта неспособность предотвратить проникновение микробов в их пористую ткань может быть основной причиной того, что у них никогда не развилась более сложная анатомия. [22]
Подобно книдариям (медузам и др.) и гребневикам (гребневикам), и в отличие от всех других известных многоклеточных животных, тела губок состоят из неживой желеобразной массы ( мезогила ), зажатой между двумя основными слоями клеток. [23] [24] Книдарии и гребневики имеют простую нервную систему, а их клеточные слои связаны внутренними связями и закреплены на базальной мембране (тонкий волокнистый мат, также известный как « базальная пластинка »). [24] У губок нет нервной системы, похожей на нервную систему позвоночных, но она может иметь совершенно иную нервную систему. [7] Их средние желеобразные слои содержат большие и разнообразные популяции клеток, а некоторые типы клеток их внешних слоев могут перемещаться в средний слой и менять свои функции. [21]
Тело губки полое и удерживает форму мезогилом , желеобразным веществом, состоящим в основном из коллагена и укрепленным плотной сетью волокон, также состоящих из коллагена. Внутренняя поверхность покрыта хоаноцитами — клетками с цилиндрическими или коническими воротниками, окружающими один жгутик на каждый хоаноцит. Волнообразные движения хлыстовидных жгутиков прогоняют воду через тело губки. У всех губок есть отверстия , каналы, ведущие внутрь через мезохил, и у большинства губок они контролируются трубчатыми пороцитами , которые образуют закрывающиеся впускные клапаны. Пинакоциты , пластинчатые клетки, образуют однослойную внешнюю оболочку над всеми остальными частями мезогиля, не покрытыми хоаноцитами, причем пинакоциты также переваривают частицы пищи, слишком крупные для попадания в отверстия, [21] [23] в то время как те, кто находится у основания животного, отвечают за его фиксацию. [23]
Внутри мезоила живут и перемещаются другие типы клеток: [21] [23]
Многие личиночные губки обладают глазами без нейронов , основанными на криптохромах . Они опосредуют фототаксическое поведение. [27]
Стеклянные губки представляют собой отличительную вариацию этого основного плана. Их спикулы, состоящие из кремнезема , образуют каркас , похожий на каркас, между стержнями которого, как паутина , подвешена живая ткань , содержащая большинство типов клеток. [21] Эта ткань представляет собой синцитий , который в некотором смысле ведет себя как множество клеток с одной внешней мембраной , а в других — как одна клетка с несколькими ядрами . Мезохил отсутствует или минимален. Цитоплазма синцития , жидкая жидкость, заполняющая внутренности клеток, организована в «реки», которые транспортируют ядра, органеллы («органы» внутри клеток) и другие вещества. [29] Вместо хоаноцитов у них есть дополнительные синцитии, известные как хоаносинцитии, которые образуют колоколообразные камеры, куда вода поступает через перфорации. Внутренняя часть этих камер выстлана «воротниковыми телами», каждое из которых состоит из воротника и жгутика, но не имеет собственного ядра. Движение жгутиков всасывает воду через ходы «паутины» и выбрасывает ее через открытые концы колоколообразных камер. [21]
Некоторые типы клеток имеют по одному ядру и мембране, но соединены с другими одноядерными клетками и с основным синцитием «мостами» из цитоплазмы . Склероциты , строящие спикулы, имеют множественные ядра , а у личинок стеклянных губок они соединены с другими тканями цитоплазматическими мостиками; такие связи между склероцитами у взрослых пока не обнаружены, но это может просто отражать трудность исследования таких мелкомасштабных особенностей. Мосты контролируются «закупоренными соединениями», которые, очевидно, пропускают одни вещества, блокируя другие. [29]
Большинство губок работают подобно дымоходам : они впитывают воду внизу и выбрасывают ее из оскулюма («маленького рта») вверху. Поскольку окружающие токи наверху быстрее, эффект всасывания, который они производят по принципу Бернулли, выполняет часть работы бесплатно. Губки могут контролировать поток воды, полностью или частично закрывая оскулум и отверстия (всасывающие поры) и изменяя ритм жгутиков, а также могут перекрыть его, если в воде много песка или ила. [21]
Хотя слои пинакоцитов и хоаноцитов напоминают эпителий более сложных животных, они не связаны прочно межклеточными связями или базальной пластинкой (тонким волокнистым листком под ней). Гибкость этих слоев и ремоделирование мезоила лофоцитами позволяют животным корректировать свою форму на протяжении всей жизни, чтобы максимально использовать преимущества местных водных потоков. [31]
Простейшая структура тела губок представляет собой трубку или вазу, известную как «асконоид», но это сильно ограничивает размер животного. Строение тела характеризуется стебельчатым спонгоцелем, окруженным одним слоем хоаноцитов. Если его просто увеличить, отношение его объема к площади поверхности увеличится, потому что поверхность увеличивается пропорционально квадрату длины или ширины, а объем увеличивается пропорционально кубу. Количество тканей, нуждающихся в питании и кислороде, определяется объемом, но насосная способность, обеспечивающая снабжение питанием и кислородом, зависит от площади, покрытой хоаноцитами. Асконоидные губки редко превышают 1 мм (0,039 дюйма) в диаметре. [21]

Некоторые губки преодолевают это ограничение, принимая «сиконоидную» структуру, при которой стенки тела складчатые . Внутренние карманы складок выстланы хоаноцитами, которые соединяются с наружными карманами складок отверстиями. Это увеличение количества хоаноцитов и, следовательно, насосной способности позволяет сиконоидным губкам вырасти до нескольких сантиметров в диаметре.
«Лейконоидная» структура еще больше увеличивает насосную способность за счет почти полного заполнения внутренней части мезогилом, который содержит сеть камер, выстланных хоаноцитами и соединенных друг с другом, а также с водозаборными и выпускными отверстиями трубками. Лейконидные губки вырастают до диаметра более 1 м (3,3 фута), а тот факт, что рост в любом направлении увеличивает количество камер хоаноцитов, позволяет им принимать более широкий диапазон форм, например, «инкрустирующие» губки, формы которых повторяют формы поверхности, к которым они прикрепляются. Все пресноводные и большинство мелководных морских губок имеют лейконидные тела. Сети водных путей стеклянных губок сходны со структурой лейконидов. [21] Во всех трех типах структуры площадь поперечного сечения областей, выстланных хоаноцитами, намного больше, чем площадь поперечного сечения впускных и выпускных каналов. Это замедляет поток вблизи хоаноцитов и, таким образом, облегчает им захват частиц пищи. [21] Например, у Leuconia , небольшой лейконоидной губки высотой около 10 сантиметров (3,9 дюйма) и диаметром 1 сантиметр (0,39 дюйма), вода поступает в каждый из более чем 80 000 всасывающих каналов со скоростью 6 см в минуту . Однако, поскольку Leuconia имеет более 2 миллионов жгутиковых камер, общий диаметр которых намного превышает диаметр каналов, скорость потока воды через камеры замедляется до 3,6 см в час , что позволяет хоаноцитам легко захватывать пищу. Вся вода выбрасывается через единственную капсулу со скоростью около 8,5 см в секунду , что достаточно быстро, чтобы уносить отходы на некоторое расстояние. [32]

В зоологии скелетом называют любую достаточно жесткую структуру животного, независимо от наличия у нее суставов и независимо от того, является ли она биоминерализованной . Мезохил у большинства губок функционирует как эндоскелет и является единственным скелетом мягких губок, который покрывает твердые поверхности, такие как камни. Чаще всего мезохил укрепляется минеральными спикулами , спонгиновыми волокнами или тем и другим. Спикулы, которые присутствуют у большинства, но не у всех видов, [33] могут быть изготовлены из кремнезема или карбоната кальция и различаются по форме от простых палочек до трехмерных «звезд» с числом лучей до шести. Спикулы производятся клетками склероцитами [21] и могут быть отдельными, соединенными суставами или слитыми. [20]
Некоторые губки также выделяют экзоскелеты , которые полностью лежат вне их органических компонентов. Например, склероспонги («твердые губки») имеют массивный экзоскелет из карбоната кальция, над которым органическое вещество образует тонкий слой с камерами хоаноцитов в ямках минерала. Эти экзоскелеты секретируются пинакоцитами , образующими кожу животных. [21]

Хотя взрослые губки в основном сидячие животные, некоторые морские и пресноводные виды могут перемещаться по морскому дну со скоростью 1–4 мм (0,039–0,157 дюйма) в день в результате амебоподобных движений пинакоцитов и других клеток. Некоторые виды могут сжимать все тело, а многие могут закрывать оскулы и отверстия. Молодые особи дрейфуют или плавают свободно, а взрослые особи неподвижны. [21]

У губок нет четко выраженной кровеносной , дыхательной , пищеварительной и выделительной систем — вместо этого все эти функции поддерживает система потока воды. Они отфильтровывают частицы пищи из протекающей через них воды. Частицы размером более 50 микрометров не могут попасть в устья, и пинакоциты поглощают их путем фагоцитоза (поглощение и внутриклеточное переваривание). Частицы размером от 0,5 мкм до 50 мкм задерживаются в отверстиях, которые сужаются от внешнего конца к внутреннему. Эти частицы поглощаются пинакоцитами или археоцитами , которые частично выдавливаются через стенки отверстий. Частицы размером с бактерии (менее 0,5 микрометра) проходят через отверстия, улавливаются и потребляются хоаноцитами . [21] Поскольку мельчайшие частицы являются наиболее распространенными, хоаноциты обычно захватывают 80% запасов пищи губки. [34] Археоциты транспортируют пищу, упакованную в пузырьки, от клеток, которые непосредственно переваривают пищу, к тем, которые этого не делают. По крайней мере, у одного вида губок есть внутренние волокна, которые функционируют как дорожки для археоцитов, переносящих питательные вещества, [21] и эти дорожки также перемещают инертные объекты. [23]
Раньше утверждалось, что стеклянные губки могут питаться питательными веществами, растворенными в морской воде, и очень не любят ила. [35] Однако исследование 2007 года не обнаружило никаких доказательств этого и пришло к выводу, что они очень эффективно (около 79%) извлекают бактерии и другие микроорганизмы из воды и перерабатывают взвешенные зерна отложений для извлечения такой добычи. [36] Воротниковые тела переваривают пищу и распределяют ее, завернутую в пузырьки, которые транспортируются «моторными» молекулами динеина по пучкам микротрубочек , которые проходят по всему синцитию . [21]
Клетки губок поглощают кислород путем диффузии из воды в клетки, когда вода течет через тело, в которую также диффундируют углекислый газ и другие растворимые отходы, такие как аммиак . Археоциты удаляют минеральные частицы, которые угрожают закупорить устья, транспортируют их через мезохил и обычно сбрасывают в выходящий поток воды, хотя некоторые виды включают их в свой скелет. [21]

В водах, где запасы пищевых частиц очень скудны, некоторые виды охотятся на ракообразных и других мелких животных. На данный момент обнаружено всего 137 видов. [38] Большинство из них принадлежат к семейству Cladorhizidae , но некоторые представители Guitarridae и Esperiopsidae также являются хищниками. [39] В большинстве случаев мало что известно о том, как они на самом деле ловят добычу, хотя считается, что некоторые виды используют либо липкие нити, либо крючковидные спикулы . [39] [40] Большинство плотоядных губок живут в глубоких водах, до 8840 м (5,49 миль), [41] и развитие методов глубоководных исследований океана, как ожидается, приведет к открытию еще нескольких. [21] [39] Однако один вид был обнаружен в средиземноморских пещерах на глубине 17–23 м (56–75 футов), наряду с более обычными губками -фильтраторами . Пещерные хищники ловят ракообразных длиной менее 1 мм (0,039 дюйма), опутывая их тонкими нитями, переваривают их, обволакивая дополнительными нитями, в течение нескольких дней, а затем возвращаются к своей нормальной форме; нет никаких доказательств того, что они используют яд . [41]
Большинство известных хищных губок полностью утратили водную систему и хоаноциты . Однако род Chondrocladia использует сильно модифицированную систему потока воды для надувания воздушных шаров, которые используются для захвата добычи. [39] [42]
Пресноводные губки часто содержат зеленые водоросли в качестве эндосимбионтов внутри археоцитов и других клеток и извлекают пользу из питательных веществ, вырабатываемых водорослями. Многие морские виды являются хозяевами других фотосинтезирующих организмов, чаще всего цианобактерий , но в некоторых случаях и динофлагеллят . Симбиотические цианобактерии могут составлять треть общей массы живой ткани некоторых губок, а некоторые губки получают от 48% до 80% своего энергоснабжения за счет этих микроорганизмов. [21] В 2008 году команда из Штутгартского университета сообщила, что спикулы из кремнезема проводят свет в мезогиль , где живут фотосинтезирующие эндосимбионты. [43] Губки, в которых обитают фотосинтезирующие организмы, наиболее распространены в водах с относительно бедным запасом пищевых частиц и часто имеют листовидную форму, которая максимизирует количество солнечного света, которое они собирают. [23]
Недавно обнаруженная плотоядная губка, обитающая вблизи гидротермальных источников, содержит бактерии , питающиеся метаном , и переваривает некоторые из них. [23]
Губки не обладают такой сложной иммунной системой, как большинство других животных. Однако они отвергают прививки от других видов, но принимают их от других представителей своего вида. У некоторых морских видов ведущую роль в отторжении чужеродного материала играют серые клетки. При проникновении они производят химическое вещество, которое останавливает движение других клеток в пораженной области, тем самым не позволяя злоумышленнику использовать внутренние транспортные системы губки. Если вторжение сохраняется, серые клетки концентрируются в этой области и выделяют токсины, которые убивают все клетки в этой области. «Иммунная» система может оставаться в этом активированном состоянии до трех недель. [23]

У губок есть три бесполых способа размножения: после фрагментации, почкованием и образованием геммул . Фрагменты губок могут быть отделены течением или волнами. Они используют подвижность своих пинакоцитов и хоаноцитов , а также изменение формы мезогиля , чтобы повторно прикрепиться к подходящей поверхности, а затем в течение нескольких дней восстанавливать себя как маленькие, но функциональные губки. Те же возможности позволяют регенерировать губкам, пропущенным через тонкую ткань. [44] Фрагмент губки может регенерировать только в том случае, если он содержит как колциниты, производящие мезохил , так и археоциты , производящие все остальные типы клеток. [34] Очень немногие виды размножаются почкованием. [45]
Геммулы — это «стручки выживания», которые несколько морских губок и многие пресноводные виды производят тысячами после отмирания и которые некоторые, в основном пресноводные виды, регулярно производят осенью. Губки образуют геммулы, окутывая оболочки спонгина, часто укрепленные спикулами, круглыми скоплениями археоцитов , полными питательных веществ. [46] Пресноводные геммулы могут также включать фотосинтезирующие симбионты. [47] Затем геммулы переходят в спячку и в этом состоянии могут пережить холод, высыхание, недостаток кислорода и резкие перепады солености . [21] Пресноводные геммулы часто не оживают до тех пор, пока температура не упадет, остаются холодными в течение нескольких месяцев, а затем достигают почти «нормального» уровня. [47] Когда геммулы прорастают, археоциты вокруг кластера снаружи превращаются в пинакоциты , мембрана над порой в оболочке лопается, кластер клеток медленно появляется, и большая часть оставшихся археоцитов трансформируется в другие типы клеток, необходимые для сделать функционирующую губку. Геммулы одного вида, но разных особей могут объединять силы, образуя одну губку. [48] Некоторые геммулы сохраняются внутри родительской губки, и весной может быть трудно определить, возродилась ли старая губка или была «реколонизирована» собственными геммулами. [47]
Большинство губок являются гермафродитами (функционируют как представители обоих полов одновременно), хотя у губок нет половых желез (органов размножения). Сперматозоиды производятся хоаноцитами или целыми камерами хоаноцитов, которые погружаются в мезохил и образуют сперматические кисты , тогда как яйцеклетки образуются путем трансформации археоцитов или хоаноцитов у некоторых видов. Каждое яйцо обычно приобретает желток , потребляя «медсестринские клетки». Во время нереста сперматозоиды вырываются из кист и выбрасываются через оскулюм . Если они контактируют с другой губкой того же вида, поток воды переносит их к хоаноцитам, которые их поглощают, но вместо переваривания превращаются в амебоидную форму и переносят сперматозоиды через мезохил к яйцеклеткам, которые в большинстве случаев поглощают носитель и свой груз. [49]
Некоторые виды выпускают оплодотворенную икру в воду, но большинство сохраняет икру до тех пор, пока она не вылупится. Существует четыре типа личинок, но все они представляют собой шарики клеток с внешним слоем клеток, жгутики или реснички которых позволяют личинкам двигаться. После плавания в течение нескольких дней личинки тонут и ползут, пока не найдут место для поселения. Большинство клеток трансформируются в археоциты, а затем в типы, соответствующие их местоположению в миниатюрной взрослой губке. [49]
Эмбрионы стеклянной губки начинают с деления на отдельные клетки, но как только образуются 32 клетки, они быстро трансформируются в личинки, которые снаружи имеют яйцевидную форму с полосой ресничек вокруг середины, которую они используют для движения, но внутри имеют типичную структуру спикул стеклянной губки с вокруг них и между ними расположен паутинистый главный синцитий , а также хоаносинцитии с многочисленными воротниковыми телами в центре. Затем личинки покидают тела своих родителей. [50]
Губки в регионах с умеренным климатом живут максимум несколько лет, но некоторые тропические виды и, возможно, некоторые глубоководные виды могут жить 200 и более лет. Некоторые кальцинированные демоспонги растут всего на 0,2 мм (0,0079 дюйма) в год, и, если эта скорость постоянна, возраст экземпляров шириной 1 м (3,3 фута) должен составлять около 5000 лет. Некоторые губки начинают половое размножение уже в возрасте нескольких недель, а другие ждут, пока им исполнится несколько лет. [21]
У взрослых губок отсутствуют нейроны или какая-либо другая нервная ткань . Однако большинство видов обладают способностью выполнять движения, скоординированные по всему телу, в основном сокращения пинакоцитов , сжимающие водные каналы и таким образом удаляющие излишки осадка и других веществ, которые могут вызвать закупорку. Некоторые виды могут сокращать оскулум независимо от остального тела. Губки также могут сокращаться, чтобы уменьшить площадь, уязвимую для нападения хищников. В тех случаях, когда две губки сливаются, например, если имеется большой, но еще не отделившийся бутон, эти волны сокращения постепенно скоординируются у обоих « сиамских близнецов ». Координирующий механизм неизвестен, но может включать химические вещества, подобные нейротрансмиттерам . [51] Однако стеклянные губки быстро передают электрические импульсы через все части синцития и используют это, чтобы остановить движение своих жгутиков , если поступающая вода содержит токсины или чрезмерный осадок. [21] Считается, что миоциты отвечают за закрытие косточки и передачу сигналов между различными частями тела. [23]
Губки содержат гены, очень похожие на те, которые содержат «рецепт» постсинаптической плотности — важной структуры приема сигналов в нейронах всех других животных. Однако у губок эти гены активируются только в «клетках колбы», которые появляются только у личинок и могут обеспечивать некоторые сенсорные способности во время плавания личинок. Это поднимает вопрос о том, представляют ли клетки колбы предшественники настоящих нейронов или являются свидетельством того, что предки губок имели настоящие нейроны, но потеряли их, когда адаптировались к сидячему образу жизни. [52]
.jpg/1280px-Euplectella_aspergillum_(cropped).jpg)
Губки распространены по всему миру и обитают в самых разных океанических средах обитания, от полярных регионов до тропиков. [34] Большинство из них живут в тихих, чистых водах, поскольку осадки, поднятые волнами или течениями, закупоривают их поры, затрудняя питание и дыхание. [35] Наибольшее количество губок обычно обитает на твердых поверхностях, таких как камни, но некоторые губки могут прикрепляться к мягким отложениям с помощью корневидного основания. [53]
Губки более многочисленны, но менее разнообразны в водах умеренного пояса, чем в тропических водах, возможно, потому, что организмы, питающиеся губками, более многочисленны в тропических водах. [54] Стеклянные губки наиболее распространены в полярных водах и в глубинах умеренных и тропических морей, поскольку их очень пористая структура позволяет им добывать пищу из этих бедных ресурсами вод с минимальными усилиями. Демогубки и известковые губки многочисленны и разнообразны в более мелких неполярных водах. [55]
Различные классы губок обитают в разных средах обитания:
Губки с фотосинтезирующими эндосимбионтами производят до трех раз больше кислорода , чем потребляют, а также больше органических веществ, чем потребляют. Такой вклад в ресурсы их среды обитания значителен вдоль Большого Барьерного рифа Австралии , но относительно незначителен в Карибском бассейне. [34]


Многие губки сбрасывают спикулы , образуя плотный ковер глубиной несколько метров, отпугивающий иглокожих , которые в противном случае охотились бы на губки. [34] Они также производят токсины, которые препятствуют росту других сидячих организмов, таких как мшанки или морские асцидии, на них или рядом с ними, что делает губки очень эффективными конкурентами за жизненное пространство. Одним из многих примеров является агелиферин .
Некоторые виды, карибская огненная губка Tedania ignis , вызывают сильную сыпь у людей, которые с ними контактируют. [21] Черепахи и некоторые рыбы питаются в основном губками. Часто говорят, что губки создают химическую защиту от таких хищников. [21] Однако эксперименты не смогли установить взаимосвязь между токсичностью химических веществ, вырабатываемых губками, и их вкусом для рыб, что уменьшило бы полезность химической защиты в качестве сдерживающего фактора. Хищничество рыбы может даже способствовать распространению губок путем отделения их фрагментов. [23] Однако некоторые исследования показали, что рыбы отдают предпочтение губкам, не имеющим химической защиты, [56] , а другое исследование показало, что высокий уровень хищничества кораллов действительно предсказывает присутствие химически защищенных видов. [57]
Стеклянные губки не производят токсичных химикатов и живут на очень глубокой воде, где хищники редки. [35]
Губки, также известные как губчатые мухи ( Neuroptera , Sisyridae ), являются специализированными хищниками пресноводных губок. Самка откладывает икру на нависающую над водой растительность. Личинки вылупляются и падают в воду, где ищут губки, чтобы питаться. Они используют свой удлиненный ротовой аппарат, чтобы протыкать губку и всасывать находящуюся внутри жидкость. Личинки некоторых видов цепляются за поверхность губки, другие укрываются во внутренних полостях губки. Взрослые личинки покидают воду и плетут кокон, в котором окукливаются. [58]
Карибская губка из куриной печени Chondrilla nucula выделяет токсины, которые убивают коралловые полипы , позволяя губкам расти на скелетах кораллов. [21] Другие, особенно представители семейства Clionaidae , используют едкие вещества, выделяемые их археоцитами, для туннелирования в камни, кораллы и раковины мертвых моллюсков . [21] Губки могут удалять с рифов до 1 м (3,3 фута) в год, создавая видимые выемки чуть ниже уровня отлива. [34]
Карибские губки рода Aplysina страдают синдромом красных полос Aplysina. Это приводит к появлению у Aplysina одной или нескольких полос ржавого цвета, иногда с прилегающими полосами некротической ткани. Эти поражения могут полностью окружать ветви губки. Болезнь, по-видимому, заразна и поражает примерно 10 процентов A. cauliformis на рифах Багамских островов. [59] Полосы ржавого цвета вызваны цианобактерией , но неизвестно, действительно ли этот организм вызывает заболевание. [59] [60]
Помимо размещения фотосинтезирующих эндосимбионтов, [21] губки известны своим широким спектром сотрудничества с другими организмами. Относительно крупная корковая губка Lissodendoryx colombiensis чаще всего встречается на каменистых поверхностях, но распространила свой ареал на луга с морской травой , позволив себе быть окруженной или заросшей губками из морских водорослей, которые неприятны местным морским звездам и поэтому защищают Lissodendoryx от них; в свою очередь, губки из морских водорослей занимают более высокие позиции вдали от отложений морского дна. [61]
Креветки рода Synalpheus образуют колонии в губках, причем каждый вид креветок населяет разные виды губок, что делает Synalpheus одним из самых разнообразных родов ракообразных . В частности, Synalpheus regalis использует губку не только в качестве источника пищи, но и в качестве защиты от других креветок и хищников. [62] Целых 16 000 особей населяют одну губку , питаясь более крупными частицами, которые собираются на губке, когда она фильтрует океан, чтобы прокормить себя. [63] Другие ракообразные, такие как раки-отшельники, обычно имеют особый вид губок, Pseudospongosorites , которые растут на них, поскольку и губка, и краб занимают раковины брюхоногих моллюсков, пока краб и губка не перерастают панцирь, что в конечном итоге приводит к тому, что краб использует тело губки в качестве защиту вместо панциря, пока краб не найдет подходящий панцирь на замену. [64]

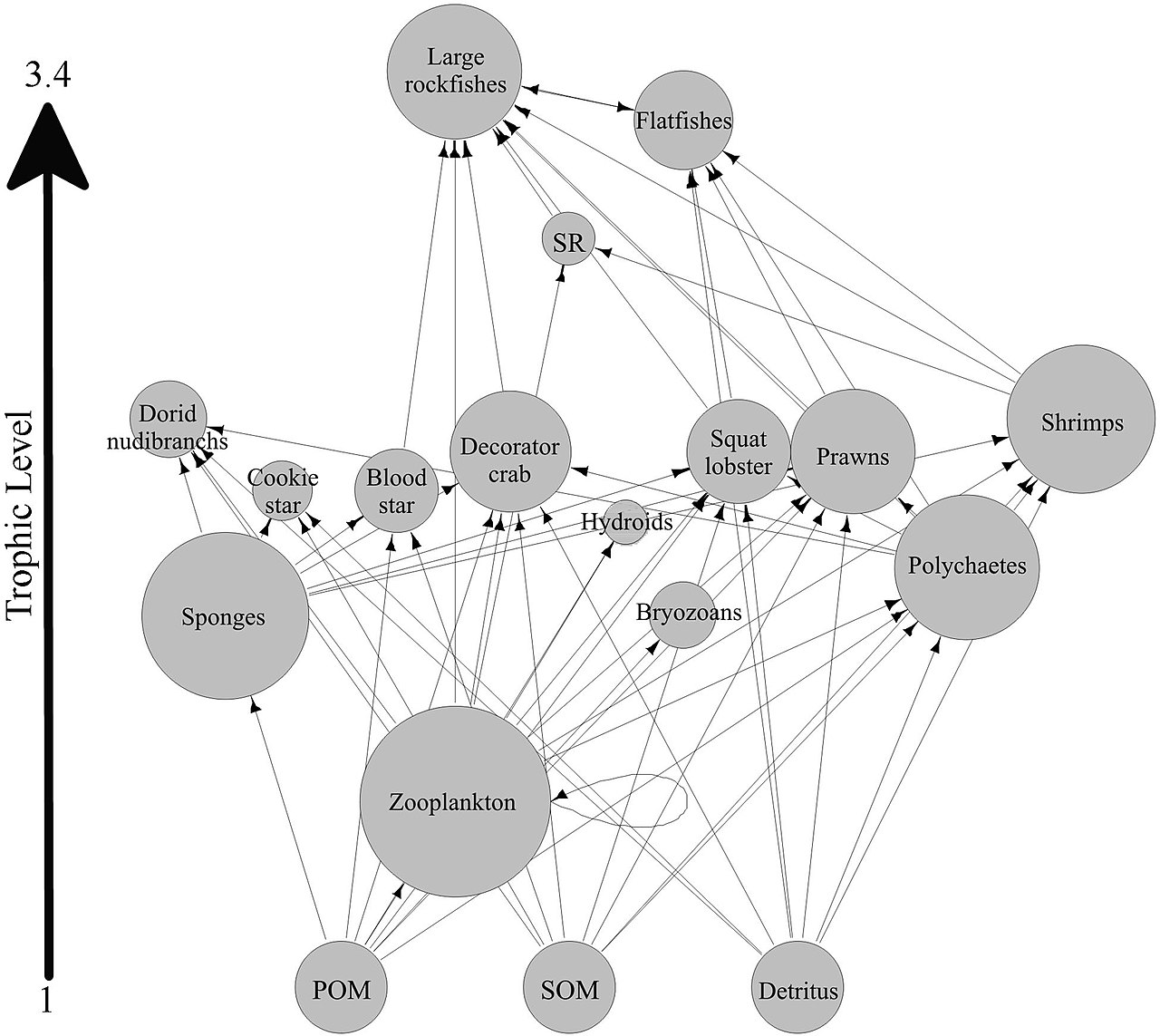
Большинство губок являются детритофагами , которые фильтруют частицы органического мусора и микроскопические формы жизни из океанской воды. В частности, губки играют важную роль в качестве детритофагов в пищевых сетях коралловых рифов , перерабатывая детрит на более высокие трофические уровни . [67]
Была выдвинута гипотеза, что губки коралловых рифов способствуют передаче органического вещества, полученного из кораллов, ассоциированным с ними детритофагам посредством производства детрита губок, как показано на диаграмме. Некоторые виды губок способны превращать полученное из кораллов РОВ в детрит губок [68] [69] и переносить органические вещества, производимые кораллами, дальше по пищевой сети рифа. Кораллы выделяют органические вещества в виде растворенной слизи и частиц, [70] [71] [72] [73] , а также клеточного материала, такого как выброшенный симбиодиниум . [74] [75] [67]
Органическое вещество может передаваться от кораллов к губкам всеми этими путями, но DOM, вероятно, составляет наибольшую долю, поскольку большая часть (от 56 до 80%) коралловой слизи растворяется в толще воды [71] и кораллы теряют фиксированный углерод. за счет изгнания симбиодиниума обычно незначительна (0,01%) [74] по сравнению с выделением слизи (до ~40%). [76] [77] Органические вещества, полученные из кораллов, также могут быть косвенно переданы губкам через бактерии, которые также могут потреблять коралловую слизь. [78] [79] [80] [67]


Помимо симбиотических отношений один к одному , хозяин может вступить в симбиоз с микробным консорциумом . Губки способны содержать широкий спектр микробных сообществ , которые также могут быть очень специфическими. Микробные сообщества, вступающие в симбиотические отношения с губкой, могут составлять до 35% биомассы ее хозяина. [84] Термин, обозначающий эти особые симбиотические отношения, при которых микробный консорциум образует пару с хозяином, называется холобиотическими отношениями . Губка, а также связанное с ней микробное сообщество будут производить широкий спектр вторичных метаболитов , которые помогают защитить ее от хищников с помощью таких механизмов, как химическая защита . [85]
Некоторые из этих взаимоотношений включают эндосимбионтов внутри клеток бактериоцитов, а также цианобактерий или микроводорослей, находящихся под слоем клеток пинакодермы, где они способны получать наибольшее количество света, используемого для фототрофии. Они могут содержать более 50 различных типов микробов и типов-кандидатов, включая Alphaprotoebacteria, Actinomycetota , Chloroflexota , Nitrospirota , « Cyanobacteria », таксоны Gamma-, тип-кандидат Poribacteria и Thaumarchaea . [85]
Линней , отнесший большинство видов сидячих животных к отряду Zoophyta класса Vermes , ошибочно отождествил род Spongia с растениями отряда Водоросли . [86] В течение долгого времени после этого губки были отнесены к отдельному подцарству Parazoa («рядом с животными»), отдельному от Eumetazoa , которое образовало остальную часть царства Animalia . [87] Их считали парафилетическим типом , из которого произошли высшие животные. [88] Другие исследования показывают, что Porifera является монофилетическим видом. [89]
Тип Porifera далее делится на классы в основном в зависимости от состава скелетов : [ 20] [34]
В 1970-х годах губки с массивным скелетом из карбоната кальция были выделены в отдельный класс Sclerospongiae , иначе известный как «кораллиновые губки». [90] Однако в 1980-х годах было обнаружено, что все они были членами либо Calcarea, либо Demospongiae. [91]
К настоящему времени в научных публикациях выявлено около 9000 видов порифер, [34] из которых: около 400 – стеклянные губки; около 500 — известковые виды; а остальные - демогубки. [21] Однако некоторые типы среды обитания, вертикальные скалы и стены пещер, а также галереи в скалах и коралловых валунах, исследованы очень мало, даже в мелководных морях. [34]
Губки традиционно делились на три класса: известковые губки (Calcarea), стеклянные губки (Hexactinellida) и демоспонгии (Demospongiae). Однако исследования показали, что Homoscleromorpha , группа, которая, как считается, принадлежит к Demospongiae , на самом деле филогенетически хорошо разделена. [92] Поэтому недавно они были признаны четвертым классом губок. [93] [94]
Губки делятся на классы в основном в зависимости от состава их скелетов : [23] Они расположены в эволюционном порядке, как показано ниже, в порядке возрастания их эволюции сверху вниз:

Хотя молекулярные часы и биомаркеры позволяют предположить, что губки существовали задолго до кембрийского взрыва жизни, спикулы кремнезема , подобные спикулам демогубок, отсутствуют в летописи окаменелостей вплоть до кембрийского периода. [96] Существует необоснованное сообщение 2002 года о спикулах в камнях, датированных примерно 750 миллионами лет назад . [97] Хорошо сохранившиеся ископаемые губки, существовавшие примерно 580 миллионов лет назад в эдиакарский период, были найдены в формации Доушантуо . Эти окаменелости, к которым относятся: спикулы; пинакоциты ; пороциты ; археоциты ; склероциты ; и внутреннюю полость, были классифицированы как демогубки. Окаменелости стеклянных губок были обнаружены примерно 540 миллионов лет назад в горных породах Австралии, Китая и Монголии. [98] Раннекембрийские губки из Мексики, принадлежащие к роду Kiwetinokia , демонстрируют свидетельства слияния нескольких более мелких спикул с образованием одной большой спикулы. [99] Спикулы карбоната кальция известковых губок были обнаружены в породах раннего кембрия примерно 530–523 миллиона лет назад в Австралии. Другие вероятные демогубки были обнаружены в фауне Чэнцзян раннего кембрия , от 525 до 520 миллионов лет назад . [100] Окаменелости, найденные на Северо-Западных территориях Канады и датируемые 890 миллионами лет назад, могут быть губками; если это открытие подтвердится, это позволит предположить, что первые животные появились до неопротерозойского события оксигенации. [101]

Пресноводные губки, по-видимому, намного моложе, поскольку самые ранние известные окаменелости датируются периодом среднего эоцена , примерно 48–40 миллионов лет назад . [98] Хотя около 90% современных губок являются демогубками , окаменелые останки этого типа встречаются реже, чем останки других типов, поскольку их скелеты состоят из относительно мягкого спонгина, который плохо окаменевает. [102] Самые ранние губчатые симбионты известны с раннего силура . [103]
Химическим индикатором является 24-изопропилхолестан , который является стабильным производным 24-изопропилхолестерина , который, как говорят, вырабатывается демоспонгами , но не эуметазойями («настоящими животными», т.е. книдариями и билатериями ). Поскольку хоанофлагелляты считаются ближайшими одноклеточными родственниками животных, группа ученых изучила биохимию и гены одного вида хоанофлагеллят . Они пришли к выводу, что этот вид не может производить 24-изопропилхолестерин, но необходимо исследование более широкого спектра хоанофлагеллят, чтобы доказать, что ископаемый 24-изопропилхолестан мог производиться только демоспонгами. [104] Хотя в предыдущей публикации сообщалось о следах химического 24-изопропилхолестана в древних породах, датируемых 1800 миллионами лет назад , [105] недавние исследования с использованием гораздо более точно датированных серий пород показали, что эти биомаркеры появляются только до конца оледенение Марино , произошедшее примерно 635 миллионов лет назад , [106] и что «анализ биомаркеров еще не выявил каких-либо убедительных доказательств существования древних губок, предшествовавших первому глобально обширному неопротерозойскому ледниковому эпизоду (Стурт, ~ 713 миллионов лет назад в Омане)» . Хотя утверждалось, что этот «биомаркер губки» мог произойти из морских водорослей, недавние исследования показывают, что способность водорослей производить этот биомаркер развилась только в каменноугольном периоде ; как таковой, биомаркер по-прежнему решительно поддерживает присутствие демоспонг в криогенном периоде. [107] [108] [109]
Археоциатиды , которые некоторые классифицируют как тип коралловой губки, являются очень распространенными окаменелостями в горных породах раннего кембрия примерно 530–520 миллионов лет назад , но, по-видимому, вымерли к концу кембрия 490 миллионов лет назад . [100] Было высказано предположение, что их производят: губки; книдарии ; водоросли ; фораминиферы ; совершенно отдельный тип животных — Archaeocyatha; или даже совершенно отдельное царство жизни, названное Archaeata или Inferibionta. С 1990-х годов археоциатиды стали рассматривать как особую группу губок. [87]
Канцеллорииды сложно вписать в классификацию губок или более сложных животных. Анализ, проведенный в 1996 году, пришел к выводу, что они были тесно связаны с губками на том основании, что детальная структура склеритов канцеллорид («броневых пластин») аналогична структуре волокон спонгина, белка коллагена , у современных кератозных (роговых) демоспонг , таких как Дарвинелла . [111] Однако другой анализ, проведенный в 2002 году, пришел к выводу, что канцеллорииды не являются губками и могут занимать промежуточное положение между губками и более сложными животными, среди прочего, потому, что их кожа была толще и более плотно соединена, чем у губок. [112] В 2008 году детальный анализ склеритов канцеллориид пришел к выводу, что они очень похожи на склериты халкиериид , мобильных двулатеральных животных, которые выглядели как слизни в кольчуге и чьи окаменелости обнаруживаются в горных породах от самого раннего кембрия до среднего кембрия. . Если это правда, то это создаст дилемму, поскольку крайне маловероятно, чтобы совершенно неродственные организмы могли независимо развить такие похожие склериты, но огромная разница в строении их тел затрудняет понимание того, как они могли быть тесно связаны. [110]

В 1990-х годах губки широко рассматривались как монофилетическая группа, все они произошли от общего предка , который сам был губкой, и как «сестринская группа» для всех других многоклеточных животных (многоклеточных животных), которые сами образуют монофилетическая группа. С другой стороны, некоторые исследования 1990-х годов также возродили идею о том, что ближайшими эволюционными родственниками животных являются хоанофлагелляты , одноклеточные организмы, очень похожие на хоаноциты губок. не быть монофилетическими, поскольку одни и те же губкообразные предки могли дать начало как современным губкам, так и негубчатым представителям Metazoa. [113]
Анализы, проведенные с 2001 года, пришли к выводу, что Eumetazoa (более сложные, чем губки) более тесно связаны с определенными группами губок, чем с другими группами губок. Такие выводы подразумевают, что губки не монофилетичны, поскольку последний общий предок всех губок также был прямым предком Eumetazoa, которые не являются губками. Исследование 2001 года, основанное на сравнении ДНК рибосом , пришло к выводу, что наиболее фундаментальное разделение внутри губок происходит между стеклянными губками и остальными, и что Eumetazoa более тесно связаны с известковыми губками (со спикулами из карбоната кальция), чем с другими типами губок. [113] В 2007 году один анализ, основанный на сравнении РНК , а другой, основанный главным образом на сравнении спикул, пришел к выводу, что демогубки и стеклянные губки более тесно связаны друг с другом, чем с известковыми губками, которые, в свою очередь, более тесно связаны с Эуметазоа. [98] [115]
Другие анатомические и биохимические данные связывают Eumetazoa с Homoscleromorpha , подгруппой демогубок. Сравнение ядерной ДНК , проведенное в 2007 году , за исключением стеклянных губок и гребневиков , показало, что:
Сперматозоиды Homoscleromorpha имеют общие черты со сперматозоидами Eumetazoa, которых нет у сперматозоидов других губок. Как у Homoscleromorpha, так и у Eumetazoa слои клеток связаны друг с другом путем прикрепления к коврообразной базальной мембране, состоящей в основном из коллагена «типа IV» , формы коллагена, не встречающейся у других губок, хотя волокна спонгина , которые укрепляют мезохил всех демогубок, аналогичен коллагену «типа IV». [25]
Описанный выше анализ пришел к выводу, что губки наиболее близки к предкам всех Metazoa, всех многоклеточных животных, включая как губок, так и более сложные группы. Однако другое сравнение, проведенное в 2008 году 150 генов в каждом из 21 рода, от грибов до человека, но включающее только два вида губок, показало, что гребневики ( гребневики ) являются самой базовой линией Metazoa, включенной в образец. [116] [117] [118] [119] Если это верно, то либо современные гребневики развили свои сложные структуры независимо от других Metazoa, либо предки губок были более сложными, и все известные губки представляют собой радикально упрощенные формы. Исследование рекомендовало провести дальнейший анализ с использованием более широкого спектра губок и других простых многоклеточных животных, таких как Placozoa . [116]
Однако повторный анализ данных показал, что компьютерные алгоритмы, использованные для анализа, были введены в заблуждение наличием специфических генов гребневиков, которые заметно отличались от таковых у других видов, в результате чего губки оставались либо сестринской группой для всех других животных, либо предковой парафилетической группой. . [120] [121] «Генеалогические древа», построенные с использованием комбинации всех доступных данных – морфологических, эволюционных и молекулярных – пришли к выводу, что губки на самом деле представляют собой монофилетическую группу, а вместе с книдариями образуют сестринскую группу билатерий. [122] [123]
Очень большое и внутренне согласованное выравнивание 1719 белков в масштабе многоклеточных животных, опубликованное в 2017 году, показало, что (i) губки, представленные Homoscleromorpha, Calcarea, Hexactinellida и Demospongiae, являются монофилетическими, (ii) губки относятся к сестринской группе для всех у других многоклеточных животных, (iii) гребневики появляются как вторая самая ранняя ветвящаяся линия животных, и (iv) плакозоя появляются как третья линия животных, за которой следуют сестринская группа книдарий и билатерий . [4]
В марте 2021 года ученые из Дублина нашли дополнительные доказательства того, что губки являются сестринской группой всех других животных, [124] а в мае 2023 года Шульц и др. обнаружили закономерности необратимых изменений в синтении генома, которые предоставляют убедительные доказательства того, что гребневики являются сестринской группой для всех других животных. [125]

В отчете 1997 года описывалось использование губок в качестве инструмента афалинами в заливе Шарк в Западной Австралии . Дельфин прикрепляет к своему роструму морскую губку , которая, по-видимому, затем используется для его защиты при поиске пищи на песчаном морском дне . [126] Поведение, известное как обтирание , наблюдалось только в этом заливе и почти исключительно проявляется самками. Исследование, проведенное в 2005 году, пришло к выводу, что матери учат этому поведению своих дочерей и что все пользователи губки тесно связаны между собой, что позволяет предположить, что это относительно недавнее нововведение. [17]


Спикулы карбоната кальция или кремнезема большинства родов губок делают их слишком грубыми для большинства применений, но два рода, Hippospongia и Spongia , имеют мягкий, полностью волокнистый скелет. [127] Ранние европейцы использовали мягкие губки для многих целей, включая прокладку для шлемов, портативную питьевую посуду и муниципальные фильтры для воды. До изобретения синтетических губок их использовали в качестве чистящих средств, аппликаторов для красок и керамической глазури, а также в качестве незаметных противозачаточных средств . Однако к середине 20-го века чрезмерный вылов рыбы привел как животных, так и эту отрасль на грань исчезновения . [128]
Многие объекты с губчатой текстурой теперь состоят из веществ, не полученных из пориферанов. К синтетическим губкам относятся личные и бытовые чистящие средства , грудные имплантаты [ 129] и противозачаточные губки . [130] Типичными используемыми материалами являются пеноцеллюлоза , пенополиуретан и реже силиконовая пена.
« Губка » люфа , также называемая люфой , которую обычно продают для использования на кухне или в душе, получена не из животного, а в основном из волокнистого «скелета» губчатой тыквы ( Luffa aegyptiaca , Cucurbitaceae ). [131]
Губки обладают лечебным потенциалом благодаря наличию в самих губках или их микробных симбионтах химических веществ, которые можно использовать для борьбы с вирусами , бактериями , опухолями и грибками. [132] [133]

Не имея какой-либо защитной оболочки или средств спасения, губки научились синтезировать множество необычных соединений. Одним из таких классов являются окисленные производные жирных кислот, называемые оксилипинами . Было обнаружено, что члены этого семейства обладают противораковыми, антибактериальными и противогрибковыми свойствами. Один из примеров, выделенный из окинавских губок плакортиса , плакоридин А , продемонстрировал потенциал цитотоксина для клеток лимфомы мышей. [134] [135]
{{cite journal}}: CS1 maint: неподходящий URL ( ссылка ){kind=link}
{kind=link}