Летучие мыши — летающие млекопитающие отряда рукокрылых ( Chiroptera , / k aɪ ˈ r ɒ p t ər ə / ). [a] Благодаря передним конечностям, приспособленным в качестве крыльев , они являются единственными млекопитающими, способными к настоящему и устойчивому полету . Летучие мыши более проворны в полете, чем большинство птиц, летая с помощью своих очень длинных растопыренных пальцев, покрытых тонкой мембраной или патагием . Самая маленькая летучая мышь и, возможно, самое маленькое из ныне живущих млекопитающих — свиноносая летучая мышь Китти , длина которой составляет 29–34 мм (1,1–1,3 дюйма), ширина крыльев — 150 мм (5,9 дюйма), а масса — 2–2,6 г (0,071–0,092 унции). Самые крупные летучие мыши — летучие лисицы , а гигантская золотистая летучая лисица ( Acerodon jubatus ) достигает веса 1,6 кг (3,5 фунта) и имеет размах крыльев 1,7 м (5 футов 7 дюймов).
Второй по величине отряд млекопитающих после грызунов , летучие мыши составляют около 20% всех классифицированных видов млекопитающих во всем мире, насчитывая более 1400 видов. Их традиционно делили на два подотряда: в основном фруктоядные мегалетучие мыши и эхолокационные микролетучие мыши . Но более поздние данные подтверждают разделение отряда на Yinpterochiroptera и Yangochiroptera , причем мегалетучие мыши являются членами первого вместе с несколькими видами микролетучих мышей. Многие летучие мыши являются насекомоядными , а большинство остальных являются фруктоядными (поедающими фрукты) или нектароядными (поедающими нектар). Несколько видов питаются животными, отличными от насекомых; например, вампирские летучие мыши питаются кровью . Большинство летучих мышей ведут ночной образ жизни , и многие из них устраиваются в пещерах или других убежищах; неясно, есть ли у летучих мышей такое поведение, чтобы спасаться от хищников . Летучие мыши присутствуют по всему миру, за исключением чрезвычайно холодных регионов. Они играют важную роль в своих экосистемах, опыляя цветы и распространяя семена; многие тропические растения в этих услугах полностью зависят от летучих мышей.
Летучие мыши приносят людям некоторые прямые выгоды ценой некоторых недостатков. Летучие мышиный помет добывался как гуано в пещерах и использовался в качестве удобрения. Летучие мыши поедают насекомых-вредителей, что снижает потребность в пестицидах и других мерах борьбы с насекомыми. Иногда их бывает достаточно много и они находятся достаточно близко к населенным пунктам, чтобы служить туристическими достопримечательностями, и их используют в пищу по всей Азии и Тихоокеанскому региону . Однако плодовые летучие мыши часто считаются вредителями садоводами. Благодаря своей физиологии летучие мыши являются одним из видов животных, которые действуют как естественный резервуар многих патогенов , таких как бешенство ; и поскольку они очень подвижны, общительны и долгоживущи, они могут легко распространять болезни между собой. Если люди взаимодействуют с летучими мышами, эти черты становятся потенциально опасными для людей. Некоторые летучие мыши также являются хищниками комаров , подавляя передачу болезней, переносимых комарами .
В зависимости от культуры летучие мыши могут символически ассоциироваться с положительными чертами, такими как защита от определенных болезней или рисков, перерождение или долгая жизнь, но на Западе летучие мыши обычно ассоциируются с тьмой, злобой, колдовством, вампирами и смертью.
Более старое английское название летучих мышей — flittermouse, что соответствует их названию в других германских языках (например, немецком Fledermaus и шведском fladdermus ), связанному с порханием крыльев. В среднеанглийском языке было bakke , скорее всего, родственно древнешведскому natbakka ( ' night-bat ' ), которое, возможно, претерпело сдвиг с -k- на -t- (в современное английское bat ) под влиянием латинского blatta , ' моль, ночное насекомое ' . Слово bat , вероятно, впервые было использовано в начале 1570-х годов. [2] [3] Название Chiroptera происходит от древнегреческого : χείρ — cheir , ' рука ' [4] и πτερόν — pteron , ' крыло ' . [1] [5]

Нежные скелеты летучих мышей плохо окаменевают; по оценкам, только 12% родов летучих мышей , которые жили, были обнаружены в палеонтологической летописи. [6] Большинство древнейших известных ископаемых летучих мышей уже были очень похожи на современных микробов, таких как Archaeopteropus (32 миллиона лет назад). Древнейшие известные ископаемые летучие мыши включают Archaeonycteris praecursor и Altaynycteris aurora (55-56 миллионов лет назад), оба известны только по изолированным зубам. [7] [8] Древнейший полный скелет летучей мыши - Icaronycteris gunnelli (52 миллиона лет назад), известный по двум скелетам, обнаруженным в Вайоминге. [9] [10] Вымершие летучие мыши Palaeochiropteryx tupaiodon и Hassianycteris kumari , оба из которых жили 48 миллионов лет назад, являются первыми ископаемыми млекопитающими, чья окраска была обнаружена: оба были красновато-коричневыми. [11] [12]
Летучие мыши ранее были сгруппированы в надотряд Archonta , вместе с древолазами (Scandentia), шерстокрылами (Dermoptera) и приматами . [13] Современные генетические данные теперь помещают летучих мышей в надотряд Laurasiatheria , с его родственным таксоном Ferungulata , который включает хищных , панголинов , непарнокопытных , парнокопытных и китообразных . [14] [15] [16] [17] [18] Одно исследование помещает рукокрылых в качестве родственного таксона для непарнокопытных (Perissodactyla). [19]
Гипотеза летающих приматов предполагает, что если убрать приспособления к полету, то мегакрылые летучие мыши становятся родственными приматам по анатомическим признакам, не свойственным микрокрылым летучим мышам, и, таким образом, полет у млекопитающих развился дважды. [20] Генетические исследования убедительно подтверждают монофилию летучих мышей и единое происхождение полета млекопитающих. [9] [20]
Независимый молекулярный анализ, пытающийся установить даты, когда эволюционировали эктопаразиты летучих мышей ( клопы ), пришел к выводу, что клопы, похожие на тех, которые известны сегодня (все основные существующие линии, все из которых питаются в основном летучими мышами), уже разнообразились и укоренились более 100 миллионов лет назад (т. е. задолго до самых старых записей о летучих мышах, 52 миллиона лет назад), предполагая, что изначально все они эволюционировали на хозяевах, не являющихся летучими мышами, и «летучие мыши были колонизированы несколько раз независимо, если только эволюционное происхождение летучих мышей не было сильно недооценено». [21] Блохи , как группа, довольно старые (большинство семейств блох образовались около конца мелового периода [22] ), но ни один анализ не предоставил оценок возраста линий блох, связанных с летучими мышами. Однако самые старые известные члены другой линии эктопаразитов летучих мышей ( летучие мухи ), появились примерно 20 миллионов лет назад, намного позже происхождения летучих мышей. [23] Семейство уховерток Arixeniidae, эктопаразитических летучих мышей , не имеет ископаемых остатков, но считается, что оно возникло не ранее 23 миллионов лет назад. [24]
Генетические данные указывают на то, что летучие мыши появились в раннем эоцене и принадлежат к четырем основным линиям летучих мышей. [18] Было предложено два новых подотряда; Yinpterochiroptera включает семейство Pteropodidae , или мегалетучих мышей, а также семейства Rhinolophidae , Hipposideridae , Craseonycteridae , Megadermatidae и Rhinopomatidae . [26] Yangochiroptera включает другие семейства летучих мышей (все из которых используют гортанную эхолокацию), вывод, подтвержденный исследованием ДНК 2005 года. [26] Филогеномное исследование 2013 года подтвердило два новых предложенных подотряда. [18]

Открытие в 2003 году ранней ископаемой летучей мыши из формации Грин-Ривер возрастом 52 миллиона лет , Onychonycteris finneyi , указывает на то, что полет развился до появления эхолокационных способностей. [27] [28] У Onychonycteris были когти на всех пяти пальцах, тогда как у современных летучих мышей максимум два когтя на двух пальцах каждой руки. У него также были более длинные задние ноги и более короткие предплечья, похожие на таковые у лазающих млекопитающих, которые висят под ветвями, таких как ленивцы и гиббоны . У этой летучей мыши размером с ладонь были короткие, широкие крылья, что говорит о том, что она не могла летать так быстро или так далеко, как более поздние виды летучих мышей. Вместо того чтобы непрерывно хлопать крыльями во время полета, Onychonycteris , вероятно, чередовал взмахи и скольжение в воздухе. [9] Это говорит о том, что эта летучая мышь не летала так много, как современные летучие мыши, а перелетала с дерева на дерево и проводила большую часть времени, карабкаясь или вися на ветвях. [29] Отличительные черты окаменелости Onychonycteris также подтверждают гипотезу о том, что полет млекопитающих, скорее всего, развился у древесных локомоторов, а не у наземных бегунов. Эта модель развития полета, обычно известная как теория «деревьев вниз», утверждает, что летучие мыши сначала летали, используя преимущество высоты и гравитации, чтобы спуститься на добычу, а не бегая достаточно быстро для взлета с уровня земли. [30] [31]
Молекулярная филогения была спорной, так как она указывала на то, что у микробов нет уникального общего предка , что подразумевало, что произошли некоторые, казалось бы, маловероятные преобразования. Первое заключается в том, что ларингеальная эхолокация дважды развилась у летучих мышей, один раз у Yangochiroptera и один раз у Rhinolophoids. [33] Второе заключается в том, что ларингеальная эхолокация имела единое происхождение у Chiroptera, впоследствии была утрачена в семействе Pteropodidae (все мегакрылые), а позже развилась как система щелкания языком в роде Rousettus . [34] Анализы последовательности гена вокализации FoxP2 не дали окончательных результатов относительно того, была ли ларингеальная эхолокация утрачена у птероподид или приобретена в эхолокационных линиях. [35] Эхолокация, вероятно, впервые произошла у летучих мышей от коммуникативных звуков. Эоценовые летучие мыши Icaronycteris (52 миллиона лет назад) и Palaeochiropteryx имели краниальные адаптации, предполагающие способность обнаруживать ультразвук . Это могло использоваться сначала в основном для поиска насекомых на земле и картирования своего окружения в фазе планирования или в коммуникативных целях. После того, как была установлена адаптация полета, она могла быть усовершенствована для нацеливания на летающую добычу с помощью эхолокации. [29] Анализ гена слуха Prestin, похоже, поддерживает идею о том, что эхолокация развивалась независимо по крайней мере дважды, а не была вторично утрачена у птероподид, [36] но онтогенетический анализ улитки подтверждает, что гортанная эхолокация развилась только один раз. [37]
Летучие мыши — плацентарные млекопитающие . После грызунов они являются крупнейшим отрядом , составляя около 20% видов млекопитающих. [38] В 1758 году Карл Линней отнес семь известных ему видов летучих мышей к роду Vespertilio в отряд Приматы . Примерно двадцать лет спустя немецкий натуралист Иоганн Фридрих Блюменбах дал им собственный отряд — Рукокрылые. [39] С тех пор число описанных видов возросло до более чем 1400, [40] традиционно классифицируемых как два подотряда: Megachiroptera (мегакрылые) и Microchiroptera (микрокрылые/эхолокационные летучие мыши). [41] Не все мегакрылые крупнее микробов. [42] Эти две группы различаются по нескольким характеристикам. Микрокрылые используют эхолокацию для навигации и поиска добычи, но мегакрылые, за исключением тех, что относятся к роду Rousettus, этого не делают. [43] Соответственно, мегалетучие мыши обладают хорошо развитым зрением. [41] У мегалетучих мышей есть коготь на втором пальце передней конечности. [44] [45] Внешние уши микролетучих мышей не закрываются, образуя кольцо; края отделены друг от друга у основания уха. [45] Мегалетучие мыши питаются фруктами , нектаром или пыльцой, в то время как большинство микролетучих мышей питаются насекомыми ; другие питаются фруктами, нектаром, пыльцой, рыбой , лягушками, мелкими млекопитающими или кровью . [41]

Ниже представлена таблица, отражающая классификацию семейств летучих мышей, признанную различными авторами девятого тома « Справочника млекопитающих мира», опубликованного в 2019 году: [46]

Форма головы и зубов летучих мышей может различаться в зависимости от вида. В целом, у мегалетучих мышей более длинные морды, большие глазницы и меньшие уши, что придает им более собачий вид, что является источником их прозвища «летучие лисицы». [47] Среди микролетучих мышей более длинные морды связаны с питанием нектаром, [48] в то время как у вампирских летучих мышей морды уменьшены для размещения больших резцов и клыков. [49]
У мелких летучих мышей, питающихся насекомыми, может быть до 38 зубов, в то время как у вампиров их всего 20. У летучих мышей, питающихся насекомыми с твердым панцирем, меньше зубов, но они крупнее, с более длинными клыками и более прочной нижней челюстью, чем у видов, питающихся насекомыми с более мягким телом. У летучих мышей, питающихся нектаром, клыки длинные, а щечные зубы уменьшены. У летучих мышей, питающихся фруктами, бугорки щечных зубов приспособлены для дробления. [48] Верхние резцы вампиров не имеют эмали , что делает их острыми как бритва. [49] Сила укуса мелких летучих мышей создается за счет механического преимущества , что позволяет им прокусывать закаленную броню насекомых или кожицу фруктов. [50]
Летучие мыши — единственные млекопитающие, способные к длительному полету, в отличие от планирования , как у белки-летяги . [51] Самая быстрая летучая мышь, мексиканская летучая мышь со свободным хвостом ( Tadarida brasiliensis ), может развивать скорость по земле до 160 км/ч (100 миль/ч). [52]
Кости пальцев летучих мышей гораздо более гибкие, чем у других млекопитающих, благодаря их уплощенному поперечному сечению и низкому уровню кальция вблизи их кончиков. [53] [54] Удлинение пальцев летучих мышей, ключевая особенность, необходимая для развития крыльев, обусловлено повышением регуляции костных морфогенетических белков (Bmps). Во время эмбрионального развития ген, контролирующий сигнал Bmp, Bmp2 , подвергается повышенной экспрессии в передних конечностях летучих мышей, что приводит к удлинению пальцев рук. Это важное генетическое изменение помогает создать специализированные конечности, необходимые для мощного полета. Относительная пропорция сохранившихся пальцев передних конечностей летучих мышей по сравнению с таковыми у ископаемых летучих мышей эоцена не имеет существенных различий, что позволяет предположить, что морфология крыльев летучих мышей сохранялась на протяжении более пятидесяти миллионов лет. [ 55] Во время полета кости подвергаются изгибу и сдвигу ; ощущаемые изгибающие напряжения меньше, чем у наземных млекопитающих, но сдвигающие напряжения больше. Кости крыльев летучих мышей имеют немного более низкую точку разрыва, чем у птиц. [56]
Как и у других млекопитающих, и в отличие от птиц, радиус является основным компонентом предплечья. У летучих мышей пять удлиненных пальцев, которые все расходятся вокруг запястья. Большой палец указывает вперед и поддерживает переднюю кромку крыла, а другие пальцы поддерживают натяжение, удерживаемое в мембране крыла. Второй и третий пальцы идут вдоль кончика крыла, позволяя крылу тянуться вперед против аэродинамического сопротивления , без необходимости быть толстыми, как у крыльев птерозавров . Четвертый и пятый пальцы идут от запястья к задней кромке и отталкивают изгибающую силу, вызванную тем, что воздух толкает жесткую мембрану. [57] Благодаря своим гибким суставам летучие мыши более маневренны и ловки, чем планирующие млекопитающие. [58]

Крылья летучих мышей намного тоньше и состоят из большего количества костей, чем крылья птиц, что позволяет летучим мышам маневрировать точнее, чем последние, и летать с большей подъемной силой и меньшим сопротивлением. [59] Складывая крылья к телу при взмахе вверх, они экономят 35 процентов энергии во время полета. [60] Мембраны нежные, легко рвутся, [61] но могут отрастать заново, и небольшие разрывы быстро заживают. [61] [62] Поверхность крыльев оснащена чувствительными к прикосновениям рецепторами на небольших выпуклостях, называемых клетками Меркеля , которые также находятся на кончиках пальцев человека. Эти чувствительные области отличаются у летучих мышей, так как каждая выпуклость имеет крошечный волосок в центре, что делает ее еще более чувствительной и позволяет летучей мыши обнаруживать и адаптироваться к изменению воздушного потока; основное использование — оценка наиболее эффективной скорости полета, а также, возможно, избежание сваливания . [63] Насекомоядные летучие мыши также могут использовать тактильные волоски, чтобы выполнять сложные маневры для захвата добычи в полете. [58]
Патагиум — это мембрана крыла; она натянута между костями руки и пальцев, а также вниз по бокам тела к задним конечностям и хвосту. Эта кожная мембрана состоит из соединительной ткани , эластичных волокон , нервов , мышц и кровеносных сосудов . Мышцы поддерживают мембрану в натянутом состоянии во время полета. [ 64] Степень, в которой хвост летучей мыши прикреплен к патагиуму, может варьироваться в зависимости от вида, при этом некоторые имеют полностью свободные хвосты или даже не имеют хвостов. [48] Кожа на теле летучей мыши, которая имеет один слой эпидермиса и дермы , а также волосяные фолликулы , потовые железы и жировой подкожный слой, сильно отличается от кожи мембраны крыла. В зависимости от вида летучей мыши наличие волосяных фолликулов и потовых желез будет варьироваться в патагиуме. [65] Этот патагиум представляет собой чрезвычайно тонкий двойной слой эпидермиса; эти слои разделены соединительнотканным центром, богатым коллагеновыми и эластичными волокнами . У некоторых видов летучих мышей потовые железы будут присутствовать между этой соединительной тканью. [66] Кроме того, если присутствуют волосяные фолликулы, это поддерживает летучую мышь, чтобы приспосабливаться к внезапным маневрам полета. [67] [68] У эмбрионов летучих мышей апоптоз (запрограммированная гибель клеток) затрагивает только задние конечности, в то время как передние конечности сохраняют перепонки между пальцами, которые формируют мембраны крыльев. [69] В отличие от птиц, чьи жесткие крылья обеспечивают изгиб и скручивание плеч, у летучих мышей гибкая мембрана крыла, которая может противостоять только растяжению. Чтобы достичь полета, летучая мышь прилагает силу внутрь в точках, где мембрана встречается со скелетом, так что противодействующая сила уравновешивает ее на краях крыла перпендикулярно поверхности крыла. Эта адаптация не позволяет летучим мышам уменьшать размах крыльев, в отличие от птиц, которые могут частично складывать крылья в полете, радикально уменьшая размах крыла и площадь для взмаха вверх и для скольжения. Следовательно, летучие мыши не могут путешествовать на большие расстояния, как птицы. [57]
Летучие мыши, питающиеся нектаром и пыльцой, могут зависать в воздухе, подобно колибри . Острые передние края крыльев могут создавать вихри , которые обеспечивают подъемную силу . Вихрь может стабилизироваться за счет изменения изгибов крыльев животным. [70]

Когда летучие мыши не летают, они висят вниз головой на своих ногах, в позе, известной как насест. [71] Бедренные кости прикреплены к бедрам таким образом, что позволяют им сгибаться наружу и вверх в полете. Голеностопный сустав может сгибаться, позволяя задней кромке крыльев сгибаться вниз. Это не позволяет совершать многие движения, кроме как висеть или карабкаться по деревьям. [57] Большинство мегалетучих мышей спят, прижав голову к животу, тогда как большинство микролетучих мышей спят, загнув шею к спине. Это различие отражено в структуре шейных или шейных позвонков в двух группах, которые четко различаются. [71] Сухожилия позволяют летучим мышам смыкать свои ноги, когда они висят на насесте. Мышечная сила нужна, чтобы отпустить, но не для того, чтобы схватить насест или держаться за него. [72]
Находясь на земле, большинство летучих мышей могут только неуклюже ползать. Несколько видов, таких как новозеландская малая короткохвостая летучая мышь и обыкновенная вампирская летучая мышь, проворны на земле. Оба вида совершают боковые походки (конечности двигаются одна за другой), когда двигаются медленно, но вампирские летучие мыши двигаются скачущей походкой (все конечности двигаются синхронно) на большей скорости, используя сложенные крылья для продвижения вперед. Летучие мыши-вампиры, вероятно, развили эти походки, чтобы следовать за своими хозяевами, в то время как короткохвостые летучие мыши развились в отсутствие конкурентов среди наземных млекопитающих. Улучшенное наземное передвижение, по-видимому, не уменьшило их способность летать. [73]
У летучих мышей эффективная кровеносная система . Похоже, они используют особенно сильную веномоцию, ритмичное сокращение мышц венозных стенок. У большинства млекопитающих стенки вен обеспечивают в основном пассивное сопротивление, сохраняя свою форму, когда через них течет дезоксигенированная кровь, но у летучих мышей они, по-видимому, активно поддерживают обратный поток крови к сердцу с помощью этого насосного действия. [74] [75] Поскольку их тела относительно небольшие и легкие, летучие мыши не подвергаются риску притока крови к их головам во время ночевки. [76]
Летучие мыши обладают высокоадаптированной дыхательной системой , чтобы справляться с требованиями мощного полета, энергетически обременительной деятельностью, которая требует большого непрерывного потока кислорода. У летучих мышей относительная площадь альвеолярной поверхности и объем легочной капиллярной крови больше, чем у большинства других мелких четвероногих млекопитающих. [77] Во время полета дыхательный цикл имеет соотношение один к одному с циклом взмахов крыльев. [78] Из-за ограничений легких млекопитающих летучие мыши не могут поддерживать полет на большой высоте. [57]
.jpg/1280px-Flying_fox_at_botanical_gardens_in_Sydney_(cropped).jpg)
Для работы летательных мышц летучих мышей требуется много энергии и эффективная кровеносная система. Энергоснабжение мышц, задействованных в полете, требует примерно вдвое больше по сравнению с мышцами, которые не используют полет как средство передвижения млекопитающих. Параллельно с потреблением энергии, уровень кислорода в крови летающих животных в два раза выше, чем у их наземных передвигающихся млекопитающих. Поскольку кровоснабжение контролирует количество кислорода, поставляемого по всему телу, кровеносная система должна реагировать соответствующим образом. Поэтому, по сравнению с наземным млекопитающим того же относительного размера, сердце летучей мыши может быть в три раза больше и перекачивать больше крови. [80] Сердечный выброс напрямую зависит от частоты сердечных сокращений и ударного объема крови; [81] активная летучая мышь может достигать частоты сердечных сокращений 1000 ударов в минуту . [82]
Благодаря своей чрезвычайно тонкой мембранной ткани крыло летучей мыши может вносить значительный вклад в общую эффективность газообмена организма. [83] Из-за высокой потребности в энергии для полета тело летучей мыши удовлетворяет эти потребности путем обмена газом через патагий крыла. Когда летучая мышь расправляет крылья, это позволяет увеличить отношение площади поверхности к объему. Площадь поверхности крыльев составляет около 85% от общей площади поверхности тела, что предполагает возможность полезной степени газообмена. [83] Подкожные сосуды в мембране лежат очень близко к поверхности и обеспечивают диффузию кислорода и углекислого газа. [84]
Пищеварительная система летучих мышей имеет различные адаптации в зависимости от вида летучей мыши и ее рациона. Как и у других летающих животных, пища обрабатывается быстро и эффективно, чтобы соответствовать потребностям в энергии. Насекомоядные летучие мыши могут иметь определенные пищеварительные ферменты для лучшей переработки насекомых, такие как хитиназа для расщепления хитина , который является большим компонентом насекомых. [85] Летучие мыши-вампиры, вероятно, из-за своей диеты, состоящей из крови, являются единственными позвоночными, у которых нет фермента мальтазы , который расщепляет солодовый сахар , в их желудочно-кишечном тракте. Нектоядные и плодоядные летучие мыши имеют больше ферментов мальтазы и сахаразы , чем насекомоядные, чтобы справиться с более высоким содержанием сахара в своем рационе. [86]
Адаптации почек летучих мышей различаются в зависимости от их рациона. Плотоядные и вампировые летучие мыши потребляют большое количество белка и могут выделять концентрированную мочу ; их почки имеют тонкую кору и длинные почечные сосочки . Плодоядные летучие мыши лишены этой способности и имеют почки, адаптированные для удержания электролитов из-за их низкоэлектролитной диеты; их почки, соответственно, имеют толстую кору и очень короткие конические сосочки. [86] У летучих мышей более высокие показатели метаболизма, связанные с полетом, что приводит к повышенной потере воды при дыхании. Их большие крылья состоят из сильно васкуляризированных мембран, что увеличивает площадь поверхности и приводит к потере воды через испарение через кожу . [79] Вода помогает поддерживать ионный баланс в их крови, системе терморегуляции и удалению отходов и токсинов из организма через мочу. Они также подвержены отравлению мочевиной крови , если не получают достаточно жидкости. [87]
Строение маточной системы у самок летучих мышей может различаться в зависимости от вида: у некоторых из них имеется два рога матки , а у других — одна основная камера. [88]
Микрокрылые и несколько мегакрылых издают ультразвуковые звуки для создания эха. Интенсивность звука этих эхо зависит от подсвязочного давления. Крикотироидная мышца летучих мышей контролирует частоту импульсов ориентации, что является важной функцией. Эта мышца расположена внутри гортани, и это единственная тензорная мышца, способная помогать фонации. [89] Сравнивая исходящий импульс с возвращающимся эхом, летучие мыши могут собирать информацию об окружающей среде. Это позволяет им обнаруживать добычу в темноте. [90] Некоторые крики летучих мышей могут достигать 140 децибел . [91] Микрокрылые используют свою гортань для испускания эхолокационных сигналов через рот или нос. [92] Крики микрокрылых имеют частоту от 14 000 до более чем 100 000 Гц, выходя далеко за пределы диапазона человеческого слуха (от 20 до 20 000 Гц). [93] У различных групп летучих мышей развились мясистые выросты вокруг и над ноздрями, известные как носовые листья , которые играют роль в передаче звука. [94]
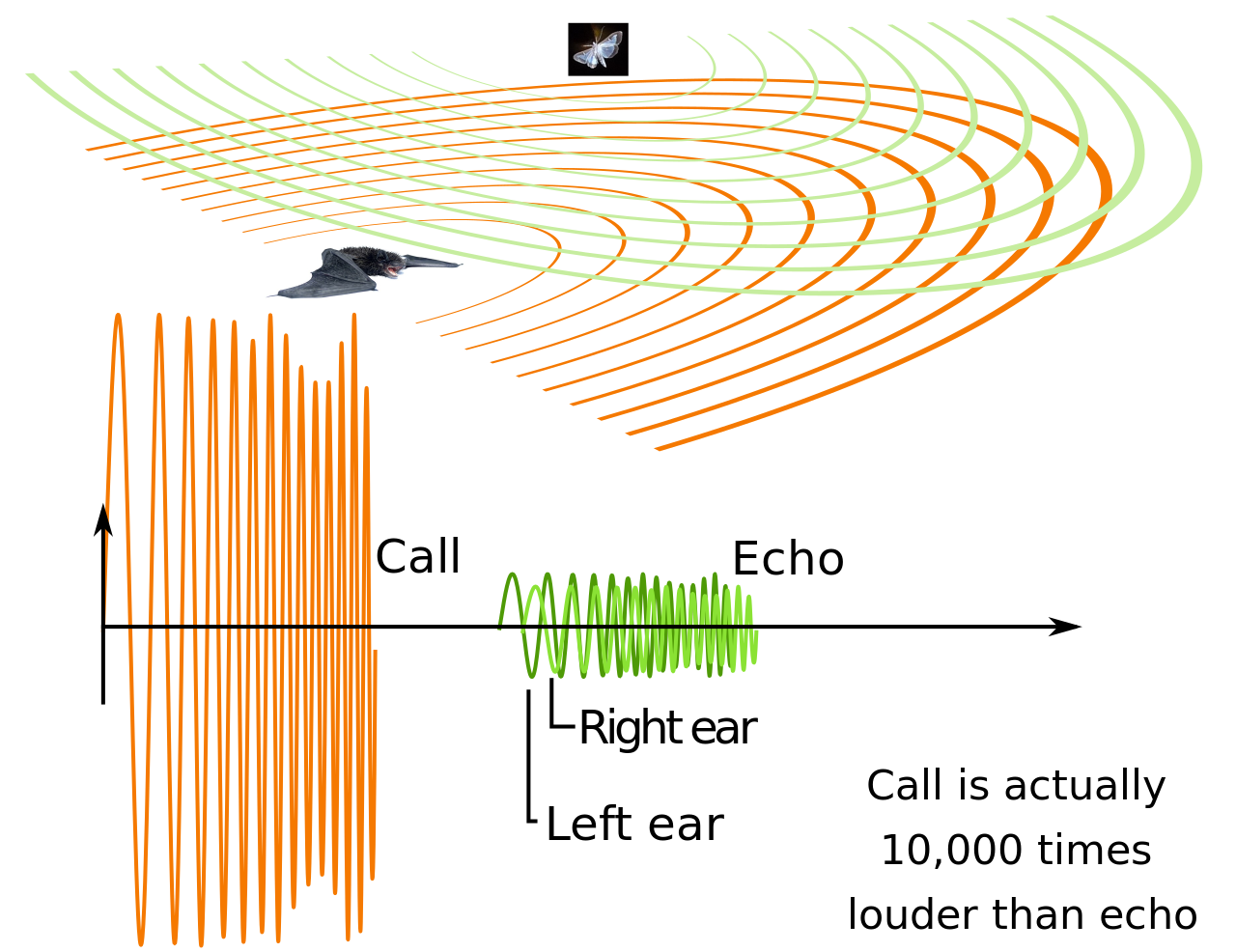
В низкочастотной эхолокации летучие мыши могут разделять свои крики и возвращающиеся эхо по времени. Им приходится рассчитывать время для своих коротких криков, чтобы они закончились до того, как вернется эхо. [95] Задержка возвращающихся эхо позволяет летучей мыши оценить расстояние до своей добычи. [93] В высокочастотной эхолокации летучие мыши издают непрерывный крик и разделяют импульс и эхо по частоте, используя эффект Доплера своего движения в полете. Сдвиг возвращающихся эхо дает информацию, касающуюся движения и местоположения добычи летучей мыши. Эти летучие мыши должны иметь дело с изменениями в доплеровском сдвиге из-за изменений скорости полета. Они приспособились изменять частоту излучения импульсов в зависимости от скорости полета, поэтому эхо по-прежнему возвращается в оптимальном диапазоне слышимости. [95] [96]
Помимо эхолокации добычи, уши летучих мышей чувствительны к звукам, издаваемым их добычей, таким как трепетание крыльев моли. Сложная геометрия гребней на внутренней поверхности ушей летучих мышей помогает четко фокусировать сигналы эхолокации и пассивно прислушиваться к любым другим звукам, издаваемым добычей. Эти гребни можно рассматривать как акустический эквивалент линзы Френеля , и они существуют у большого количества неродственных животных, таких как айе -айе , малый галаго , большеухая лисица , мышиный лемур и другие. [97] [98] [99] Летучие мыши могут оценивать высоту своей цели, используя интерференционные картины от эха, отражающегося от козелка , лоскута кожи во внешнем ухе. [93]

С помощью повторного сканирования летучие мыши могут мысленно создавать точное изображение окружающей среды, в которой они движутся, и своей добычи. [102] Некоторые виды моли используют это, например, тигровая моль , которая производит апосематические ультразвуковые сигналы, чтобы предупредить летучих мышей о том, что они химически защищены и поэтому неприятны на вкус. [100] [101] Виды моли, включая тигровую моль, могут производить сигналы, чтобы подавить эхолокацию летучих мышей . У многих видов моли есть орган слуха, называемый тимпаном , который реагирует на входящий сигнал летучей мыши, заставляя летательные мышцы моли хаотично подергиваться, отправляя моль в случайные маневры уклонения. [103] [104] [105]
Глаза большинства видов микробов маленькие и плохо развиты, что приводит к плохой остроте зрения , но ни один вид не слеп. [106] Большинство микробов имеют мезопическое зрение , что означает, что они могут обнаруживать свет только на низких уровнях, тогда как другие млекопитающие имеют фотопическое зрение , которое обеспечивает цветное зрение. Микролетучие мыши могут использовать свое зрение для ориентации и во время перемещения между местами ночлега и кормления, поскольку эхолокация эффективна только на коротких расстояниях. Некоторые виды могут обнаруживать ультрафиолет (УФ). Поскольку тела некоторых микробов имеют отчетливую окраску, они могут различать цвета. [51] [107] [108] [109]
Виды мегабатов часто имеют зрение, такое же хорошее, как человеческое, если не лучше. Их зрение адаптировано как к ночному, так и к дневному зрению, включая некоторое цветовое зрение. [109]
Микролетучие мыши используют магниторецепцию , поскольку они обладают высокой чувствительностью к магнитному полю Земли , как и птицы. Микролетучие мыши используют компас, основанный на полярности, что означает, что они различают север и юг, в отличие от птиц, которые используют силу магнитного поля для различения широт , что может быть использовано при дальних путешествиях. Механизм неизвестен, но может включать частицы магнетита . [110] [111]

Большинство летучих мышей являются гомойотермными (имеют стабильную температуру тела), исключение составляют летучие мыши-веперы (Vespertilionidae), подковоносые летучие мыши (Rhinolophidae), свободнохвостые летучие мыши (Molossidae) и летучие мыши с изогнутыми крыльями (Miniopteridae), которые широко используют гетеротермию (когда температура тела может меняться). [112] [113] По сравнению с другими млекопитающими летучие мыши обладают высокой теплопроводностью . Крылья заполнены кровеносными сосудами и теряют тепло тела при вытягивании. В состоянии покоя они могут обвивать свои крылья вокруг себя, чтобы удерживать слой теплого воздуха. У более мелких летучих мышей, как правило, более высокая скорость метаболизма, чем у более крупных, и поэтому им нужно потреблять больше пищи, чтобы поддерживать гомойотермию. [114]
Летучие мыши могут избегать полетов в течение дня, чтобы предотвратить перегрев на солнце, так как их темные мембраны крыльев поглощают солнечное излучение. Летучие мыши могут быть не в состоянии рассеивать тепло, если температура окружающей среды слишком высока; [115] они используют слюну, чтобы охладиться в экстремальных условиях. [57] Среди мегалетучих мышей летучая лисица Pteropus hypomelanus использует слюну и взмахи крыльев, чтобы охладиться во время ночевки в самое жаркое время дня. [116] Среди микролетучих мышей юмская нетопырь ( Myotis yumanensis ), мексиканская свободнохвостая летучая мышь и бледная летучая мышь ( Antrozous pallidus ) справляются с температурами до 45 °C (113 °F), тяжело дыша, выделяя слюну и облизывая свой мех, чтобы способствовать испарительному охлаждению; этого достаточно, чтобы рассеивать вдвое больше вырабатываемого ими метаболического тепла. [117]
У летучих мышей также есть система сфинктерных клапанов на артериальной стороне сосудистой сети, которая проходит вдоль края их крыльев. Когда они полностью открыты, они позволяют насыщенной кислородом крови течь через капиллярную сеть через мембрану крыла; когда они сжимаются, они направляют поток непосредственно в вены, минуя капилляры крыла. Это позволяет летучим мышам контролировать, сколько тепла обменивается через мембрану полета, позволяя им выделять тепло во время полета. Многие другие млекопитающие используют капиллярную сеть в больших ушах для той же цели. [118]

Оцепенение , состояние пониженной активности, при котором температура тела и метаболизм снижаются, особенно полезно для летучих мышей, поскольку они используют большое количество энергии во время активности, зависят от ненадежного источника пищи и имеют ограниченную способность запасать жир. Обычно они понижают температуру своего тела в этом состоянии до 6–30 °C (43–86 °F) и могут сократить свои энергетические затраты на 50–99%. [119] Тропические летучие мыши могут использовать его, чтобы избежать хищников, сокращая время, затрачиваемое на поиски пищи, и тем самым уменьшая вероятность быть пойманными хищником. [120] Обычно считалось, что мегабаты гомойотермны, но три вида небольших мегабатов, массой около 50 граммов ( 1+3 ⁄ 4 унций), как известно, используют оцепенение: обыкновенный цветочный летучий мышь ( Syconycteris australis ), длинноязыкий нектарник ( Macroglossus minimus ) и восточный трубконосый летучий мышь ( Nyctimene robinsoni ). Оцепеневшие состояния длятся дольше летом у мегалетучих мышей, чем зимой. [121]
Во время спячки летучие мыши впадают в спячку и снижают температуру своего тела на 99,6% периода спячки; даже в периоды возбуждения, когда они возвращают свою температуру тела к нормальной, они иногда впадают в неглубокое спячку, известное как «гетеротермическое возбуждение». [122] Некоторые летучие мыши впадают в спячку при более высоких температурах , чтобы сохранять прохладу в летние месяцы. [123]
Гетеротермные летучие мыши во время длительных миграций могут летать ночью и впадать в торпидное состояние, ночуя днем. В отличие от перелетных птиц, которые летают днем и питаются ночью, у ночных летучих мышей есть конфликт между путешествием и едой. Сэкономленная энергия снижает их потребность в пище, а также сокращает продолжительность миграции, что может помешать им проводить слишком много времени в незнакомых местах и уменьшить хищничество. У некоторых видов беременные особи могут не использовать торпор. [124] [125]
Самая маленькая летучая мышь — свиноносая летучая мышь Китти ( Craseonycteris thonglongyai ), длина которой составляет 29–34 мм ( 1+1 ⁄ 8 – 1+3 ⁄ 8 дюйма) в длину с размахом крыльев 150 мм (6 дюймов) и весом2–2,6 г ( 1 ⁄ 16 – 3 ⁄ 32 унции). [126] Это также, возможно, самый маленький из ныне живущих видов млекопитающих, после этрусской землеройки . [127] Самые крупные летучие мыши — это несколько видов крыланов Pteropus megabats и гигантская летучая лисица с золотой короной ( Acerodon jubatus ), которая может весить1,6 кг ( 3+1 ⁄ 2 фунта) с размахом крыльев 1,7 м (5 футов 7 дюймов). [128] Более крупные летучие мыши, как правило, используют более низкие частоты, а более мелкие — более высокие для эхолокации; высокочастотная эхолокация лучше подходит для обнаружения более мелкой добычи. Мелкая добыча может отсутствовать в рационе крупных летучих мышей, поскольку они не способны ее обнаружить. [129] Адаптации конкретного вида летучих мышей могут напрямую влиять на то, какие виды добычи им доступны. [130]

Полет позволил летучим мышам стать одной из наиболее широко распространенных групп млекопитающих. [131] За исключением Арктики, Антарктики и нескольких изолированных океанических островов, летучие мыши существуют практически в каждой среде обитания на Земле. [132] В тропических зонах, как правило, обитает больше видов, чем в умеренных. [133] Различные виды выбирают разные среды обитания в разные сезоны, от побережий до гор и пустынь, но им требуются подходящие места для ночлега. Гнезда летучих мышей можно найти в дуплах, расщелинах, листве и даже в созданных человеком сооружениях, и включают в себя «палатки», которые летучие мыши строят из листьев. [134] Мегалетучие мыши обычно ночуют на деревьях. [135] Большинство микролетучих мышей ведут ночной образ жизни [136] , а мегалетучие мыши, как правило, ведут дневной или сумеречный образ жизни . [137] [138] Известно, что летучие мыши ведут дневной образ жизни в умеренных регионах летом, когда ночью недостаточно времени для поиска пищи, [139] [140] и в районах, где днем мало пернатых хищников. [141] [142]
В умеренных зонах некоторые микролетучие мыши мигрируют на сотни километров в зимние спячки; [143] другие впадают в оцепенение в холодную погоду, пробуждаясь и питаясь, когда теплая погода позволяет насекомым быть активными. [144] Другие уходят в пещеры на зиму и впадают в спячку на срок до шести месяцев. [144] Микролетучие мыши редко летают в дождь; он мешает их эхолокации, и они не могут охотиться. [145]
Различные виды летучих мышей имеют разный рацион питания, включая насекомых, нектар, пыльцу, фрукты и даже позвоночных. [146] Мегабаты в основном питаются фруктами, нектаром и пыльцой. [137] Из-за своего небольшого размера, высокого метаболизма и быстрого сжигания энергии во время полета летучие мыши должны потреблять большое количество пищи для своего размера. Насекомоядные летучие мыши могут съедать более 120 процентов своего веса в день, в то время как плодоядные летучие мыши могут съедать более чем в два раза больше своего веса. [147] Они могут путешествовать на значительные расстояния каждую ночь, в исключительных случаях до 38,5 км (24 мили) у пятнистой летучей мыши ( Euderma maculatum ) в поисках пищи. [148] Летучие мыши используют различные стратегии охоты. [129] Летучие мыши получают большую часть своей воды из пищи, которую они едят; многие виды также пьют из водных источников, таких как озера и ручьи, летая над поверхностью и опуская язык в воду. [149]
Рукокрылые в целом находятся в процессе утраты способности синтезировать витамин С. [ 150] В ходе испытаний 34 видов летучих мышей из шести основных семейств, включая основные семейства летучих мышей, питающихся насекомыми и фруктами, было обнаружено, что все они утратили способность синтезировать его, и эта утрата может происходить от общего предка летучих мышей как единичная мутация. [151] [b] По крайней мере два вида летучих мышей, плодоядная летучая мышь ( Rousettus leschenaultii ) и насекомоядная летучая мышь ( Hipposideros armiger ), сохранили свою способность вырабатывать витамин С. [152]
Большинство летучих мышей, особенно в умеренных зонах, охотятся на насекомых. [146] Рацион насекомоядных летучих мышей может охватывать множество видов, [153] включая мух , комаров , жуков , моль, кузнечиков , сверчков , термитов , пчел , ос , поденок и ручейников . [48] [154] [155] Большое количество мексиканских свободнохвостых летучих мышей ( Tadarida brasiliensis ) летают на высоте сотен метров над землей в центральном Техасе, чтобы питаться мигрирующими молями. [156] Виды, которые охотятся на насекомых в полете, такие как маленькая коричневая летучая мышь ( Myotis lucifugus ), могут поймать насекомое в воздухе ртом и съесть его в воздухе или использовать свои хвостовые перепонки или крылья, чтобы подхватить насекомое и отнести его ко рту. [157] [158] Летучая мышь также может принести насекомое обратно в свое гнездо и съесть его там. [159] Более медленные виды летучих мышей, такие как коричневый ушан ( Plecotus auritus ) и многие виды подковоносов, могут подбирать или собирать насекомых с растительности или охотиться на них с насестов. [48] Насекомоядные летучие мыши, живущие в высоких широтах, должны потреблять добычу с более высокой энергетической ценностью, чем тропические летучие мыши. [160]

Фруктоядность, или плодоядность, встречается в обоих основных подотрядах. Летучие мыши предпочитают спелые фрукты, срывая их с деревьев зубами. Они возвращаются в свои убежища, чтобы съесть фрукты, высасывая сок и выплевывая семена и мякоть на землю. Это помогает распространять семена этих фруктовых деревьев, которые могут пускать корни и расти там, где их оставили летучие мыши, и многие виды растений зависят от летучих мышей в распространении семян . [161] [162] Ямайская фруктовая летучая мышь ( Artibeus jamaicensis ) была зарегистрирована как переносящая плоды весом 3–14 г ( 1 ⁄ 8 – 1 ⁄ 2 унции) или даже до 50 г ( 1+3 ⁄ 4 унции). [163]
Летучие мыши, питающиеся нектаром, приобрели специализированные адаптации. Эти летучие мыши обладают длинными мордами и длинными, выдвижными языками, покрытыми тонкими щетинками, которые помогают им питаться определенными цветами и растениями. [162] [164] Трубкогубая нектарница ( Anoura fistulata ) имеет самый длинный язык среди всех млекопитающих относительно размера своего тела. Это полезно для них с точки зрения опыления и питания. Их длинные, узкие языки могут проникать глубоко в длинную чашеобразную форму некоторых цветов. Когда язык втягивается, он сворачивается внутри грудной клетки. [164] Из-за этих особенностей летучие мыши, питающиеся нектаром, не могут легко перейти к другим источникам пищи в периоды дефицита, что делает их более склонными к вымиранию, чем другие виды летучих мышей. [165] [166] Питание нектаром также помогает различным растениям, поскольку эти летучие мыши служат опылителями , поскольку пыльца прикрепляется к их меху во время питания. Около 500 видов цветковых растений зависят от опыления летучими мышами и поэтому имеют тенденцию открывать свои цветы ночью. [162] Многие растения тропических лесов зависят от опыления летучими мышами. [167]

Некоторые летучие мыши охотятся на других позвоночных, таких как рыбы, лягушки, ящерицы, птицы и млекопитающие. [48] [169] Бахромчатогубая летучая мышь ( Trachops cirrhosus ), например, искусна в ловле лягушек. Эти летучие мыши обнаруживают большие группы лягушек, отслеживая их брачные крики, а затем выхватывая их с поверхности воды своими острыми клыками. [170] Большая вечерница может ловить птиц в полете. [168] Некоторые виды, такие как большая бульдоговая летучая мышь ( Noctilio leporinus ), охотятся на рыбу. Они используют эхолокацию, чтобы обнаружить небольшую рябь на поверхности воды, пикируют и используют специально увеличенные когти на задних лапах, чтобы схватить рыбу, затем относят свою добычу к месту кормления и съедают ее. [171] Известно, что по крайней мере два вида летучих мышей питаются другими летучими мышами: летучая мышь-призрак ( Vampyrum spectrum ) и летучая мышь-призрак ( Macroderma gigas ). [172]
.jpg/1280px-Desmo-boden_(cropped).jpg)
Несколько видов, в частности, обычные, белокрылые и мохнатоногие вампиры, питаются только кровью животных ( гематофагия ). Обычные вампиры обычно питаются крупными млекопитающими, такими как крупный рогатый скот ; мохнатоногие и белокрылые вампиры питаются птицами. [173] Вампиры нацеливаются на спящую добычу и могут определять глубокое дыхание. [174] Тепловые датчики в носу помогают им обнаруживать кровеносные сосуды вблизи поверхности кожи. [175] Они прокалывают кожу животного зубами, откусывая небольшой лоскут, [176] и слизывают кровь языком, который имеет боковые бороздки, приспособленные для этой цели. [177] Кровь не сворачивается благодаря антикоагулянту в слюне. [176]
Летучие мыши подвергаются нападению хищных птиц , таких как совы , ястребы и соколы , а в местах ночлега — наземных хищников, способных лазать, таких как кошки. [178] Летающие низко летучие мыши уязвимы для крокодилов . [179] Известно, что двадцать видов тропических змей Нового Света ловят летучих мышей, часто ожидая у входов в убежища, такие как пещеры, когда летучие мыши пролетят мимо. [180] Дж. Райделл и Дж. Р. Спикман утверждают, что летучие мыши перешли на ночной образ жизни в период раннего и среднего эоцена , чтобы избегать хищников. [178] Некоторые зоологи считают, что доказательства пока неоднозначны. [181]
.jpg/1280px-Little_Brown_Bat_with_White_Nose_Syndrome_(Greeley_Mine,_cropped).jpg)
Как и большинство млекопитающих, летучие мыши являются хозяевами ряда внутренних и внешних паразитов. [182] Среди эктопаразитов летучие мыши переносят блох и клещей , а также специфических паразитов, таких как летучие мыши и летучие мухи ( Nycteribiidae и Streblidae ). [183] [184] Летучие мыши являются одними из немногих отрядов неводных млекопитающих, которые не являются хозяевами вшей , возможно, из-за конкуренции со стороны более специализированных паразитов, которые занимают ту же нишу. [184]
Синдром белого носа — это состояние , связанное с гибелью миллионов летучих мышей в восточной части Соединенных Штатов и Канаде. [185] Заболевание названо в честь белого грибка Pseudogymnoascus destructans , который растет на мордах, ушах и крыльях пораженных летучих мышей. Грибок в основном передается от летучей мыши к летучей мыши и вызывает заболевание. [186] Грибок был впервые обнаружен в центральной части штата Нью-Йорк в 2006 году и быстро распространился по всей восточной части США к северу от Флориды; в большинстве пораженных пещер наблюдались показатели смертности 90–100%. [187] С 2006 года в Новой Англии и среднеатлантических штатах целые виды были полностью истреблены, а другие с численностью, которая выросла от сотен тысяч, даже миллионов, до нескольких сотен или меньше. [188] Новая Шотландия, Квебек, Онтарио и Нью-Брансуик стали свидетелями идентичных случаев вымирания, при этом канадское правительство готовится защитить все оставшиеся популяции летучих мышей на своей территории. [189] Научные данные свидетельствуют о том, что более длительные зимы, когда грибок имеет более длительный период для заражения летучих мышей, приводят к большей смертности. [190] [191] [192] В 2014 году инфекция пересекла реку Миссисипи, [193] а в 2017 году ее обнаружили у летучих мышей в Техасе. [194]
Летучие мыши являются естественными резервуарами для большого количества зоонозных патогенов , [195] включая бешенство , эндемичное для многих популяций летучих мышей, [196] [197] [198] гистоплазмоз как напрямую, так и через гуано, [199] вирусы Нипах и Хендра , [200] [201] и, возможно, вирус Эбола , [202] [203] естественный резервуар которого пока неизвестен. [204] [205] Их высокая мобильность, широкое распространение, большая продолжительность жизни, значительная симпатрия (перекрытие ареалов) видов и социальное поведение делают летучих мышей благоприятными хозяевами и переносчиками болезней. [206] Обзоры нашли разные ответы на вопрос, имеют ли летучие мыши больше зоонозных вирусов, чем другие группы млекопитающих. В одном обзоре 2015 года было обнаружено, что летучие мыши, грызуны и приматы являются носителями значительно большего количества зоонозных вирусов (которые могут передаваться человеку), чем другие группы млекопитающих, хотя различия между вышеупомянутыми тремя группами не были значительными (у летучих мышей не больше зоонозных вирусов, чем у грызунов и приматов). [207] В другом обзоре млекопитающих и птиц 2020 года было обнаружено, что идентичность таксономических групп не оказывает никакого влияния на вероятность наличия зоонозных вирусов. Вместо этого более разнообразные группы имеют большее вирусное разнообразие. [208]
Они, по-видимому, обладают высокой устойчивостью ко многим патогенам, которые они переносят, что предполагает определенную степень адаптации их иммунной системы. [206] [209] [210] Их взаимодействие с домашними животными и скотом, включая хищничество летучих мышей-вампиров, случайные встречи и поедание трупов летучих мышей, усугубляет риск зоонозной передачи. [197] Летучие мыши причастны к возникновению тяжелого острого респираторного синдрома (ТОРС) в Китае, поскольку они служат естественными хозяевами коронавирусов , несколько из которых происходят из одной пещеры в Юньнани , один из которых превратился в вирус ТОРС. [199] [211] [212] Однако они не вызывают и не распространяют COVID-19 . [213]
Некоторые летучие мыши ведут одиночный образ жизни, в то время как другие живут колониями, насчитывающими более миллиона особей. [214] Например, мексиканская летучая мышь со свободным хвостом пролетает более тысячи миль до пещеры шириной 100 футов (30 м), известной как пещера Бракен , каждый год с марта по октябрь, где обитает поразительное количество в двадцать миллионов особей этого вида, [215] тогда как летучая мышь с ушами мышей живет почти полностью одиночно. [216] Жизнь в больших колониях снижает риск нападения хищников на особь. [48] Виды летучих мышей умеренного климата могут собираться в роях в местах спячки с приближением осени. Это может служить для знакомства молодняка с местами спячки, сигналом к размножению у взрослых особей и позволяет взрослым особям размножаться с особями из других групп. [217]
У нескольких видов существует социальная структура деления-слияния , при которой большое количество летучих мышей собирается в одном месте ночевки, а также происходит разделение и смешивание подгрупп. В рамках этих сообществ летучие мыши способны поддерживать долгосрочные отношения. [218] Некоторые из этих отношений состоят из матрилинейно связанных самок и их зависимого потомства. [219] Совместное питание и взаимный уход могут происходить у некоторых видов, таких как обыкновенный вампир ( Desmodus rotundus ), и это укрепляет социальные связи. [220] [221] Гомосексуальная фелляция наблюдалась у бонинской летучей лисицы Pteropus pselaphon [222] и индийской летучей лисицы Pteropus medius [223] , хотя функция и цель этого поведения неясны.

Летучие мыши являются одними из самых голосистых млекопитающих и производят звуки, чтобы привлечь партнёров, найти партнёров для ночлега и защитить ресурсы. Эти звуки, как правило, низкочастотные и могут распространяться на большие расстояния. [48] [225] Мексиканские свободнохвостые летучие мыши являются одними из немногих видов, которые «поют» как птицы. Самцы поют, чтобы привлечь самок. Песни состоят из трёх фраз: щебетание, трель и жужжание, первые имеют слоги «A» и «B». Песни летучих мышей весьма стереотипны, но с вариациями в количестве слогов, порядке фраз и повторениях фраз между особями. [224] Среди больших копьеносых летучих мышей ( Phyllostomus hastatus ) самки производят громкие широкополосные звуки среди своих партнёров по ночлегу, чтобы сформировать групповую сплочённость. Звуки различаются между группами ночёвок и могут возникать в результате вокального обучения. [226]
В исследовании египетских крыланов, содержащихся в неволе, исследователи смогли идентифицировать 70% направленных звуков, указав, какая особь их издала, а 60% можно было разделить на четыре контекста: ссора из-за еды, толкотня из-за положения в их спящем скоплении, протест из-за попыток спаривания и спор, когда они сидели в непосредственной близости друг от друга. Животные издавали немного разные звуки при общении с разными особями летучих мышей, особенно противоположного пола. [227] У сильно диморфной по половому признаку молотоголовой летучей мыши ( Hypsignathus monstrosus ) самцы издают глубокие, резонирующие, монотонные звуки, чтобы привлечь самок. Летучие мыши в полете издают голосовые сигналы для регулирования движения. Большие бульдоги издают гудящие сигналы, когда сталкиваются друг с другом. [225]
Летучие мыши также общаются другими способами. Самцы маленьких желтоплечих летучих мышей ( Sturnira lilium ) имеют плечевые железы, которые производят пряный запах во время сезона размножения. Как и у многих других видов, у них есть волосы, специализированные для удержания и рассеивания выделений. Такие волосы образуют заметный воротник вокруг шеи некоторых самцов мегалетучих мышей Старого Света. Самцы больших мешкокрылых летучих мышей ( Saccopteryx bilineata ) имеют мешочки в крыльях, в которых они смешивают выделения тела, такие как слюна и моча, чтобы создать духи, которые они разбрызгивают на местах ночлега, поведение, известное как «соление». Соление может сопровождаться пением. [225]

Большинство видов летучих мышей являются полигамными , когда самцы спариваются с несколькими самками. Самцы нетопыря-карлика, вечерницы и вампира могут претендовать на ресурсы, которые привлекают самок, например, места ночевок, и защищать их, и спариваться с этими самками. Самцы, неспособные претендовать на место, вынуждены жить на периферии, где у них меньше репродуктивного успеха. [228] [48] Промискуитет , когда оба пола спариваются с несколькими партнерами, существует у таких видов, как мексиканская свободнохвостая летучая мышь и маленькая коричневая летучая мышь. [229] [230] По-видимому, у этих летучих мышей есть предвзятость в отношении определенных самцов среди самок. [48] У некоторых видов, таких как желтокрылая летучая мышь и спектральная летучая мышь, взрослые самцы и самки образуют моногамные пары. [48] [231] Спаривание лек , когда самцы объединяются и конкурируют за выбор самки посредством демонстрации, редко встречается у летучих мышей [232], но встречается у молотоголовой летучей мыши. [233]
У летучих мышей умеренного климата спаривание происходит в конце лета и начале осени. [234] Тропические летучие мыши могут спариваться в сухой сезон. [235] После спаривания самец может оставить после себя брачную пробку , чтобы заблокировать сперму других самцов и таким образом обеспечить свое отцовство. [236] Известно, что у видов, впадающих в спячку, самцы спариваются с самками, находящимися в оцепенении. [48] Самки летучих мышей используют различные стратегии для контроля сроков беременности и рождения детенышей, чтобы роды совпадали с максимальной пищевой способностью и другими экологическими факторами. У самок некоторых видов наблюдается отсроченное оплодотворение, при котором сперма хранится в репродуктивном тракте в течение нескольких месяцев после спаривания. Спаривание происходит в конце лета - начале осени, но оплодотворение происходит только в конце зимы - начале весны. У других видов наблюдается отсроченная имплантация , при которой яйцо оплодотворяется после спаривания, но остается свободным в репродуктивном тракте до тех пор, пока внешние условия не станут благоприятными для рождения и ухода за потомством. [237] В другой стратегии происходит и оплодотворение, и имплантация, но развитие плода задерживается до тех пор, пока не наступят хорошие условия. Во время задержанного развития мать поддерживает оплодотворенную яйцеклетку живой с помощью питательных веществ. Этот процесс может продолжаться в течение длительного периода из-за развитой системы газообмена. [238]

Для летучих мышей умеренного климата роды обычно происходят в мае или июне в Северном полушарии; роды в Южном полушарии происходят в ноябре и декабре. Тропические виды рожают в начале сезона дождей. [239] У большинства видов летучих мышей самки вынашивают и рожают одного детеныша на помет. [240] При рождении детеныш летучей мыши может весить до 40 процентов веса матери, [48] а тазовый пояс самки может расширяться во время родов, поскольку две половины соединены гибкой связкой. [241] Самки обычно рожают в положении головой вверх или горизонтально, используя силу тяжести для облегчения родов. Детеныш появляется задом вперед, возможно, чтобы предотвратить запутывание крыльев, и самка держит его в своих перепонках крыла и хвоста. У многих видов самки рожают и воспитывают детенышей в родильных колониях и могут помогать друг другу в родах. [242] [243] [241]
Большая часть заботы о молодой летучей мыши исходит от матери. У моногамных видов роль играет отец. Алло-сосание, когда самка кормит грудью детенышей другой матери, встречается у нескольких видов. Это может служить для увеличения размера колонии у видов, где самки возвращаются в свою родную колонию для размножения. [48] Способность молодой летучей мыши летать совпадает с развитием взрослого тела и длиной передних конечностей. У маленькой коричневой летучей мыши это происходит примерно через восемнадцать дней после рождения. Отлучение детенышей от груди у большинства видов происходит менее чем через восемьдесят дней. Обыкновенная летучая мышь-вампир выкармливает свое потомство дольше этого времени, и молодые летучие мыши-вампиры достигают независимости позже в жизни, чем другие виды. Вероятно, это связано с диетой этого вида, основанной на крови, которую трудно получить ночью. [244]

Максимальная продолжительность жизни летучих мышей в три с половиной раза больше, чем у других млекопитающих аналогичного размера. Было зарегистрировано шесть видов, которые живут более тридцати лет в дикой природе: бурый ушан ( Plecotus auritus ), малая бурая ночница ( Myotis lucifugus ), сибирская ночница ( Myotis sibiricus ), малая мышеухая ночница ( Myotis blythii ) , большая подковоносая летучая мышь ( Rhinolophus ferrumequinum ) и индийская летучая лисица ( Pteropus giganteus ). [245] Одна из гипотез, согласующаяся с теорией скорости жизни, связывает это с тем фактом, что они замедляют свой метаболизм во время спячки ; летучие мыши, которые впадают в спячку, в среднем имеют более длинную продолжительность жизни, чем те, которые этого не делают. [246] [247]
Другая гипотеза заключается в том, что полеты снизили их уровень смертности, что также справедливо для птиц и планирующих млекопитающих. Виды летучих мышей, которые рожают нескольких детенышей, как правило, имеют более короткую продолжительность жизни, чем виды, которые рождают только одного детеныша. Виды, гнездящиеся в пещерах, могут иметь более длинную продолжительность жизни, чем виды, не гнездящиеся в пещерах, из-за снижения хищников в пещерах. Самец сибирской летучей мыши был пойман в дикой природе через 41 год, что делает его самой старой известной летучей мышью. [247] [248]
Статус сохранности летучих мышей по состоянию на 2020 год по данным МСОП (всего 1314 видов) [249]
Такие группы, как Bat Conservation International [250], стремятся повысить осведомленность об экологической роли летучих мышей и экологических угрозах, с которыми они сталкиваются. Эта группа призвала проводить Неделю признания летучих мышей с 24 по 31 октября каждого года, чтобы повысить осведомленность об экологической важности летучих мышей. [251] В Соединенном Королевстве все летучие мыши защищены в соответствии с Законами о дикой природе и сельской местности , и потревожить летучую мышь или ее гнездо может быть наказано большим штрафом. [252] В Сараваке , Малайзия, «все летучие мыши» [253] защищены в соответствии с Указом о защите дикой природы 1998 года , [253] но такие виды, как безволосая летучая мышь ( Cheiromeles torquatus ), по-прежнему употребляются в пищу местными общинами. [254] В современной истории люди стали причиной вымирания нескольких видов летучих мышей, последним из которых стал нетопырь острова Рождества ( Pipistrellus murrayi ), который был объявлен вымершим в 2009 году. [255]
Многие люди строят домики для летучих мышей, чтобы привлечь летучих мышей. [256] Дом для летучих мышей, построенный в Университете Флориды в 1991 году, является крупнейшим в мире обитаемым искусственным убежищем, в котором проживает около 400 000 человек. [257] В Великобритании толстостенные и частично подземные доты времен Второй мировой войны были переоборудованы в убежища для летучих мышей, [258] [259] а также специально построенные дома для летучих мышей иногда строятся, чтобы смягчить ущерб среде обитания от дорог или других разработок. [260] [261] Иногда устанавливаются пещерные ворота , чтобы ограничить вход человека в пещеры с чувствительными или находящимися под угрозой исчезновения видами летучих мышей. Ворота спроектированы так, чтобы не ограничивать поток воздуха, и, таким образом, поддерживать микроэкосистему пещеры. [262] Из 47 видов летучих мышей, обнаруженных в Соединенных Штатах, 35, как известно, используют человеческие сооружения, включая здания и мосты. Четырнадцать видов используют домики для летучих мышей. [263]
Летучих мышей едят в странах Африки, Азии и Тихоокеанского региона. В некоторых случаях, например, на Гуаме, летучие лисицы оказались под угрозой исчезновения из-за охоты на них ради еды. [264] Существуют данные, свидетельствующие о том, что ветряные турбины могут создавать достаточную баротравму (повреждение давлением), чтобы убить летучих мышей. [265] У летучих мышей типичные легкие млекопитающих , которые, как считается, более чувствительны к резким изменениям давления воздуха, чем легкие птиц , что делает их более подверженными фатальному разрыву. [266] [267] [268] [269] [270] Летучих мышей могут привлекать турбины, возможно, в поисках убежища, что увеличивает уровень смертности. [266] Акустические отпугиватели могут помочь снизить смертность летучих мышей на ветряных электростанциях. [271]
Диагноз и вклад баротравмы в гибель летучих мышей вблизи лопастей ветряных турбин были оспорены другими исследованиями, сравнивавшими мертвых летучих мышей, найденных вблизи ветряных турбин, с летучими мышами, погибшими в результате столкновения со зданиями в районах, где турбин нет. [ 272]
,_from_Los_Caprichos_-_Google_Art_Project.jpg/1280px-Francisco_Jos%C3%A9_de_Goya_y_Lucientes_-_The_sleep_of_reason_produces_monsters_(No._43),_from_Los_Caprichos_-_Google_Art_Project.jpg)
Поскольку летучие мыши являются млекопитающими, но могут летать, в различных традициях они считаются пограничными существами . [273] Во многих культурах, в том числе в Европе, летучие мыши ассоциируются с тьмой, смертью, колдовством и злобой. [274] Среди коренных американцев, таких как крики , чероки и апачи , летучая мышь идентифицируется как обманщик . [275] В Танзании крылатое существо, похожее на летучую мышь, известное как Попобава, считается злым духом, меняющим форму , который нападает на своих жертв и занимается содомией . [276] В мифологии ацтеков летучие мыши символизировали землю мертвых, разрушение и упадок. [277] [278] [279] В восточно-нигерийской сказке говорится, что летучая мышь развила свои ночные привычки после того, как стала причиной смерти своего партнера, кустарниковой крысы, и теперь прячется днем, чтобы избежать ареста. [280]
В некоторых культурах существуют более позитивные изображения летучих мышей. В Китае летучие мыши ассоциировались со счастьем, радостью и удачей. Пять летучих мышей используются для символизации «Пяти благословений»: долголетия, богатства, здоровья, любви к добродетели и мирной смерти. [281] Летучая мышь священна в Тонга и часто считается физическим проявлением отделимой души . [282] В цивилизации сапотеков в Мезоамерике бог летучей мыши покровительствовал кукурузе и плодородию. [283]

Weird Sisters в шекспировском «Макбете» использовали мех летучей мыши в своем вареве. [284] В западной культуре летучая мышь часто является символом ночи и ее зловещей природы. Летучая мышь является основным животным, связанным с вымышленными персонажами ночи, как со зловещими вампирами , такими как граф Дракула и до него Варни Вампир , [285] так и с героями, такими как персонаж DC Comics Бэтмен . [286] Романы Кеннета Оппеля Silverwing повествуют о приключениях молодой летучей мыши, [287] основанной на серебристой летучей мыши Северной Америки. [288]
Летучая мышь иногда используется как геральдический символ в Испании и Франции, появляясь на гербах городов Валенсия , Пальма-де-Майорка , Фрага , Альбасете и Моншове . [289] [290] [291] Три штата США имеют официальную государственную летучую мышь . Техас и Оклахома представлены мексиканской свободнохвостой летучей мышью, в то время как Вирджиния представлена виргинской большеухой летучей мышью ( Corynorhinus townsendii virginianus ). [292]
Насекомоядные летучие мыши особенно полезны фермерам, поскольку они контролируют популяции сельскохозяйственных вредителей и снижают необходимость использования пестицидов . Было подсчитано, что летучие мыши экономят сельскохозяйственной отрасли Соединенных Штатов от 3,7 до 53 миллиардов долларов в год на пестицидах и ущербе урожаю. Это также предотвращает чрезмерное использование пестицидов, которые могут загрязнять окружающую среду и могут привести к резистентности у будущих поколений насекомых. [293]
Летучая мышь, тип гуано , богата нитратами и добывается из пещер для использования в качестве удобрения . [294] Во время Гражданской войны в США селитру собирали в пещерах для изготовления пороха . В то время считалось, что весь нитрат поступает из гуано летучих мышей, но теперь известно, что большая его часть производится нитрифицирующими бактериями . [295]
Мост Конгресс-авеню в Остине, штат Техас , является летним домом для крупнейшей в Северной Америке городской колонии летучих мышей, насчитывающей, по оценкам, 1 500 000 мексиканских свободнохвостых летучих мышей. Около 100 000 туристов в год посещают мост в сумерках, чтобы посмотреть, как летучие мыши покидают насест. [296]
Несколько молекулярных исследований показали, что рукокрылые относятся к Laurasiatheria (представленному плотоядными, панголинами, цетариодактилями, эулипотифланами и непарнокопытными) и лишь отдаленно связаны с дермоптеранами, скандентиями и приматами. (Никайдо и др., 2000 г.; Лин и Пенни, 2001 г.; Мэдсен и др., 2001 г.; Мерфи и др., 2001
г.
, 2001
г
.; Ван Ден Буше и Хуфер, 2004 г.)
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на ноябрь 2024 г. ( ссылка )A. fistulata
(на фото лакающая сахарную воду из трубки) имеет самый длинный язык по отношению к длине тела среди всех млекопитающих — и теперь ученые думают, что знают, почему.
известные в настоящее время вирусы, обнаруженные у летучих мышей, и выделены риски передачи их человеку.
Летучие мыши, которые представляют примерно 24% всех известных видов млекопитающих, часто выступают в качестве переносчиков лиссавирусов.
В этой статье мы описываем выделение HeV из летучих мышей-крылокрылых, подтверждая наши серологические и эпидемиологические доказательства того, что эти животные являются естественным резервуаром-хозяином этого вируса.
Мы обнаружили доказательства бессимптомной инфекции вирусом Эбола у трех видов летучих мышей, что указывает на то, что эти животные могут выступать в качестве резервуара для этого смертельного вируса.
Летучие мыши, давно известные как переносчики бешенства, могут быть источником некоторых из самых смертоносных новых вирусов, включая SARS, Эболу, Нипах, Хендра и Марбург.Примечание: Это краткий обзор различных научных публикаций, упомянутых в предыдущем предложении.
не знают, откуда взялся вирус Эбола.
Несмотря на согласованные усилия по расследованию, естественный резервуар вируса неизвестен.
Генетическое разнообразие последовательностей, полученных от летучих мышей, подтверждает представление о том, что летучие мыши являются естественным резервуаром-хозяином кластера коронавирусов SARS.
«Шейд основан на серебристой летучей мыши. Я думал, что они очень лихо выглядят. Мне понравилось, что эта летучая мышь жила в той же части света, что и я (восточная Канада). Это маленькие существа с размахом крыльев в несколько дюймов. Их тела примерно такого же размера, как у мышей. Они насекомоядные, что означает, что они едят только насекомых». – KO
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка )