Брюхоногие моллюски ( / ˈ ɡ æ s t r ə p ɒ d z / ) , широко известные как слизни и улитки , принадлежат к большому таксономическому классу беспозвоночных внутри типа Mollusca , называемому Gastropoda ( / ɡ æ s ˈ t r ɒ p ə d ə / ). [5]
В этот класс входят улитки и слизни, обитающие в соленой, пресной воде и на суше. Существуют многие тысячи видов морских улиток и слизней , а также пресноводных улиток , пресноводных блюдечек , наземных улиток и слизней .
Класс Gastropoda — разнообразный и весьма успешный класс моллюсков типа Mollusca. Он содержит огромное количество названных видов, уступая по общему количеству только насекомым . Ископаемая история этого класса восходит к позднему кембрию . По состоянию на 2017 год известно [обновлять]721 семейство брюхоногих моллюсков, из которых 245 вымерли и встречаются только в летописи окаменелостей , а 476 в настоящее время сохранились с летописью окаменелостей или без нее. [6]
Брюхоногие моллюски (ранее известные как одностворчатые моллюски и иногда называемые «Gasteropoda») составляют основную часть типа Mollusca и являются наиболее разнообразным классом этого типа , насчитывающим от 65 000 до 80 000 [3] [4] живых видов улиток и слизней . Анатомия , поведение, питание и репродуктивная адаптация брюхоногих значительно различаются от одной клады или группы к другой, поэтому сформулировать общие сведения для всех брюхоногих моллюсков сложно.
Класс Gastropoda отличается исключительным разнообразием ареалов . Представители обитают в садах, лесах, пустынях и горах; в небольших канавах, больших реках и озерах; в эстуариях , илистых отмелях , каменистой литорали , песчаной сублиторали, абиссальных глубинах океанов, включая гидротермальные жерла , и многочисленных других экологических нишах, в том числе паразитических .
Хотя название «улитка» может применяться и часто применяется ко всем членам этого класса, обычно это слово означает только те виды, у которых внешняя оболочка достаточно велика, чтобы мягкие части могли полностью уйти в нее. Слизни — это брюхоногие моллюски, у которых нет панциря или очень маленькая внутренняя раковина; полуслизни — это брюхоногие моллюски, у которых есть панцирь, в который они могут частично, но не полностью спрятаться.
К морским видам брюхоногих моллюсков относятся такие виды, как морское ушко , раковины , барвинки , трубачи и многие другие морские улитки, у которых ракушки сворачиваются на взрослой стадии, хотя у некоторых скручивание может быть не очень заметно, например, у каури . . У ряда семейств видов, например у всех разнообразных блюдец , панцирь скручивается только на личиночной стадии, а после этого представляет собой простую коническую структуру.
В научной литературе брюхоногие моллюски были описаны как « брюхоногие » Жоржем Кювье в 1795 году . «Нога» животного расположена ниже его кишок. [7]
Более раннее название «одностворчатый» означает одну створку (или раковину), в отличие от двустворчатых моллюсков , таких как моллюски, у которых есть две створки или раковины.
На всех таксономических уровнях брюхоногие моллюски по разнообразию уступают только насекомым . [8]
Наибольшее количество названных видов моллюсков имеют брюхоногие моллюски . Однако оценки общего количества видов брюхоногих сильно различаются в зависимости от цитируемых источников. О количестве видов брюхоногих моллюсков можно судить по оценкам числа описанных видов моллюсков с принятыми названиями: около 85 000 (минимум 50 000, максимум 120 000). [9] Но оценка общего числа моллюсков, включая неописанные виды, составляет около 240 000 видов. [10] Оценка 85 000 моллюсков включает 24 000 описанных видов наземных брюхоногих моллюсков. [9]
По разным оценкам водных брюхоногих (по разным источникам) насчитывается около 30 000 видов морских брюхоногих и около 5 000 видов пресноводных и солоноватых брюхоногих моллюсков. Многие глубоководные виды еще предстоит открыть, поскольку только 0,0001% глубоководного дна изучено биологически. [11] [12] Общее количество ныне живущих видов пресноводных улиток составляет около 4000. [13]
Недавно вымершие виды брюхоногих моллюсков (вымершие с 1500 г.) насчитывают 444 вида, 18 видов сейчас вымерли в дикой природе (но все еще существуют в неволе), а 69 видов «возможно вымерли». [14]
Число доисторических (ископаемых) видов брюхоногих моллюсков составляет не менее 15 000 видов. [15]
В морских местообитаниях наибольшее разнообразие наблюдается на континентальном склоне и континентальном поднятии , тогда как на континентальном шельфе и абиссальных глубинах разнообразие морских брюхоногих моллюсков низкое. [16]

Брюхоногие моллюски встречаются в самых разных водных и наземных средах обитания, от глубоких океанских впадин до пустынь.
Некоторые из наиболее знакомых и известных брюхоногих моллюсков — это наземные брюхоногие моллюски (наземные улитки и слизни). Некоторые обитают в пресной воде, но большинство названных видов брюхоногих обитают в морской среде.
Брюхоногие моллюски имеют широкое распространение по всему миру, от ближней Арктики и Антарктики до тропиков. Они адаптировались практически ко всем видам существования на Земле, колонизировав почти все доступные среды обитания.
В местах обитания, где недостаточно карбоната кальция для построения действительно прочного панциря, например, на некоторых кислых почвах на суше, встречаются различные виды слизней, а также некоторые улитки с тонкими полупрозрачными панцирями, большей частью или полностью состоящими из белка конхиолина .
Улитки, такие как Sphincterochila boissieri и Xerocrassaseetzeni, приспособились к условиям пустыни. Другие улитки приспособились к существованию в канавах, вблизи глубоководных гидротермальных источников , в океанических впадинах на глубине 10 000 метров под поверхностью, [17] в бурных прибоях скалистых берегов , пещерах и многих других разнообразных местах.
Брюхоногие моллюски могут быть случайно перенесены из одного места обитания в другое другими животными, например птицами . [18]
Улитки отличаются анатомическим процессом, известным как перекручивание , при котором внутренняя масса животного во время развития поворачивается на 180° в одну сторону, так что анус располагается более или менее над головой. Этот процесс не связан со скручиванием оболочки, которое представляет собой отдельное явление. Перекрут присутствует у всех брюхоногих моллюсков, но у заднежаберных брюхоногих моллюсков вторичное искривление в разной степени. [19] [20]
Перекручивание происходит в два этапа. Первая, механистическая стадия – мышечная, вторая – мутагенетическая. Эффекты перекрута в первую очередь физиологические. Организм развивается путем асимметричного роста, при этом большая часть роста происходит на левой стороне. Это приводит к утрате анатомии правой стороны, которая у большинства билатерий является дубликатом анатомии левой стороны. Существенной особенностью этой асимметрии является то, что анус обычно располагается по одну сторону от срединной плоскости. Жаберные гребешки , органы обоняния , ножная слизевая железа , нефридии и ушная раковина сердца на одной стороне тела одиночные или, по крайней мере, более развиты, чем на другой. Кроме того, имеется только одно половое отверстие, расположенное на той же стороне тела, что и анус. [21] Более того, анус перенаправляется в то же пространство, что и голова. Предполагается, что это имеет некоторую эволюционную функцию, поскольку до перекрута при втягивании в раковину сначала втягивался задний конец, а затем передний. Теперь фронт легче отвести, что, возможно, указывает на оборонительную цель.
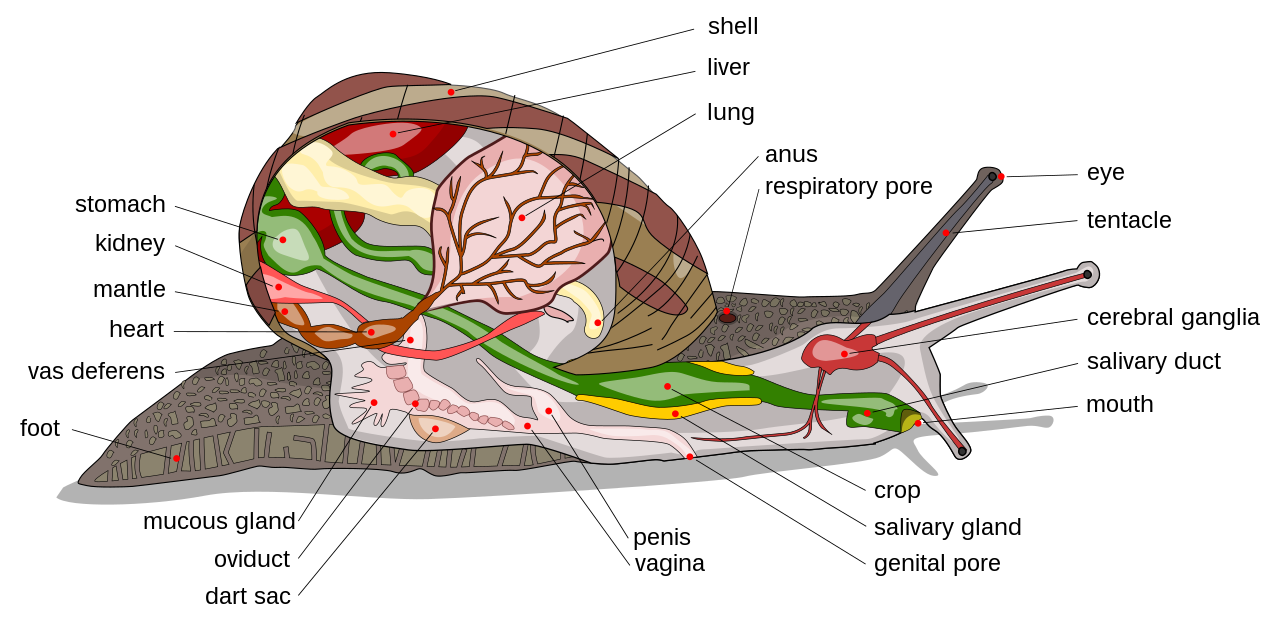
Брюхоногие моллюски обычно имеют четко выраженную голову с двумя или четырьмя сенсорными щупальцами с глазами и брюшную ногу. Передний отдел стопы называется проподиумом. Его функция — отталкивать осадок при ползании улитки. Личиночная оболочка брюхоногих моллюсков называется протоконхом .
.jpg/1280px-Zonitoides_nitidus_shell_(cropped).jpg)
Большинство брюхоногих моллюсков с панцирем имеют цельную раковину (за исключением двустворчатых брюхоногих моллюсков ), обычно свернувшуюся в спираль или спираль, по крайней мере, на личиночной стадии. Эта спиральная раковина обычно открывается с правой стороны (если смотреть вершиной раковины вверх ). У многих видов есть жаберная крышка , которая у многих видов действует как люк, закрывающий раковину. Обычно он состоит из роговидного материала, но у некоторых моллюсков он известковый. У наземных слизней панцирь редуцирован или отсутствует, тело обтекаемой формы.
У некоторых брюхоногих раков взрослые особи имеют тяжелое дно из-за наличия толстого, часто широкого, выпуклого вентрального отложения мозоли на внутренней губе, расположенного адапикально к отверстию, что может быть важно для гравитационной устойчивости. [22]
Некоторые морские слизни имеют очень яркую окраску. Это служит либо предупреждением , когда они ядовиты или содержат стрекательные клетки , либо маскировкой их на ярко окрашенных гидроидах , губках и морских водорослях, на которых обитают многие виды.
Боковые выросты на теле голожаберных называются цератами . Они содержат карманы пищеварительных желез , называемые дивертикулами .

К органам чувств брюхоногих моллюсков относятся органы обоняния , глаза, статоцисты и механорецепторы . [23] У брюхоногих нет слуха. [23]
У наземных брюхоногих моллюсков (наземных улиток и слизней) важнейшим органом чувств являются органы обоняния, расположенные на кончиках четырёх щупалец . [23] Хемосенсорные органы заднежаберных морских брюхоногих моллюсков называются ринофорами .
У большинства брюхоногих моллюсков имеются простые органы зрения — глазные пятна на кончике или основании щупалец. Однако «глаза» у брюхоногих варьируются от простых глазков , которые различают только свет и темноту, до более сложных ямочных глаз и даже глаз-хрусталиков . [24] У наземных улиток и слизней зрение не является самым важным чувством, поскольку они в основном ведут ночной образ жизни. [23]
Нервная система брюхоногих включает периферическую нервную систему и центральную нервную систему . Центральная нервная система состоит из ганглиев , соединенных нервными клетками. В него входят парные ганглии: церебральные ганглии, педальные ганглии, осфрадиальные ганглии, плевральные ганглии, париетальные ганглии и висцеральные ганглии. Иногда встречаются также буккальные ганглии. [23]
Радула брюхоногих моллюсков обычно приспособлена к пище, которую употребляет данный вид. Простейшими брюхоногими моллюсками являются блюдечки и морские ушки , травоядные животные, которые используют свою твердую радулу , чтобы скрежетать водоросли на камнях.
Многие морские брюхоногие моллюски роют норы и имеют сифон , выступающий от края мантии . Иногда в раковине имеется сифональный канал для размещения этой структуры. Сифон позволяет животному втягивать воду в мантийную полость и через жабры. Они используют сифон в первую очередь, чтобы «попробовать» воду и обнаружить добычу на расстоянии. Брюхоногие моллюски с сифонами, как правило, либо хищники, либо падальщики.
Почти все морские брюхоногие моллюски дышат жабрами , но у многих пресноводных видов и большинства наземных видов имеются мантийные легкие . Дыхательным белком почти у всех брюхоногих моллюсков является гемоцианин , но у одного пресноводного семейства легочных , Planorbidae , в качестве дыхательного белка используется гемоглобин .
У одной большой группы морских слизней жабры расположены в виде розетки перистых перьев на спине, что и дало начало их другому названию — голожаберные . У некоторых голожаберников спина гладкая или бородавчатая , без видимого жаберного механизма, поэтому дыхание, вероятно, может происходить непосредственно через кожу.
Брюхоногие моллюски имеют открытую систему кровообращения , транспортной жидкостью является гемолимфа . Гемоцианин присутствует в гемолимфе в качестве дыхательного пигмента.
Основными органами выделения у брюхоногих моллюсков являются нефридии , которые в качестве отходов выделяют либо аммиак , либо мочевую кислоту . Нефридий также играет важную роль в поддержании водного баланса у пресноводных и наземных видов. К дополнительным органам выделения, по крайней мере у некоторых видов, относятся перикардиальные железы в полости тела и пищеварительные железы, открывающиеся в желудок .

Ухаживание является частью брачного поведения у некоторых брюхоногих моллюсков, в том числе у некоторых Helicidae . Опять же, у некоторых наземных улиток необычной особенностью репродуктивной системы брюхоногих моллюсков является наличие и использование любовных стрел .
У многих морских брюхоногих моллюсков, кроме заднежаберных , имеются раздельнополые ( раздельнополые / гонохорные ); Однако большинство наземных брюхоногих моллюсков — гермафродиты .

Ухаживание является частью поведения спаривающихся брюхоногих моллюсков с некоторыми легочными семействами наземных улиток, создающих и использующих любовные дротики , метание которых было идентифицировано как форма полового отбора . [25]
К основным аспектам жизненного цикла брюхоногих моллюсков относятся:

Рацион брюхоногих моллюсков различается в зависимости от рассматриваемой группы. Морские брюхоногие моллюски включают травоядных , питающихся детритом , хищных плотоядных , падальщиков , паразитов , а также несколько ресничных питающихся, у которых радула редуцирована или отсутствует. Наземные виды могут жевать листья, кору, фрукты и разлагающихся животных, а морские виды могут соскребать водоросли с камней на морском дне. Некоторые виды, такие как Archaeogastropda, имеют горизонтальные ряды тонких краевых зубов. У некоторых видов, которые превратились в эндопаразитов, таких как эвлимида Thyonicola Doglieli , многие стандартные особенности брюхоногих моллюсков сильно редуцированы или отсутствуют.
Некоторые морские слизни являются травоядными, а некоторые — хищниками. Плотоядность обусловлена специализацией. Многие брюхоногие моллюски имеют особые пищевые предпочтения и регулярно встречаются в тесной связи со своими видами пищи.
К некоторым хищным плотоядным брюхоногим моллюскам относятся, например: Конусные ракушки , Тестацеллы , Даудебардии , Турриды , Слизни-призраки и другие.
Брюхоногие моллюски демонстрируют значительную степень вариаций в организации митохондриальных генов по сравнению с другими животными. [26] Основные события реаранжировки генов произошли у Patellogastropoda и Heterobranchia , тогда как меньшее количество изменений произошло между предками Vetigastropoda (только тРНК D, C и N) и Caenogastropoda (большая одиночная инверсия и транслокации тРНК D и N). Н). [26] Внутри Heterobranchia порядок генов кажется относительно консервативным, а реаранжировки генов в основном связаны с транспозицией генов тРНК. [26]


Первые брюхоногие моллюски были исключительно морскими, причем самые ранние представители группы появились в позднем кембрии ( Chippewaella , Strepsodiscus ), [27] однако их единственным признаком брюхоногих моллюсков является спиральный панцирь, поэтому они могли принадлежать к стволовой линии, если бы были брюхоногие моллюски вообще. [28] Самые ранние кембрийские организмы, такие как Helcionella , Barskovia и Scenella , больше не считаются брюхоногими моллюсками, а крошечная свернутая спиралью Aldanella раннего кембрийского времени, вероятно , даже не моллюск. [ нужна цитата ]
Таким образом, только в ордовике появляются первые члены коронной группы. [29] К ордовикскому периоду брюхоногие моллюски представляли собой разнообразную группу, обитающую в различных водных средах обитания. Обычно ископаемые брюхоногие моллюски из пород раннего палеозоя слишком плохо сохранились для точной идентификации. Тем не менее силурийский род Poleumita включает пятнадцать идентифицированных видов. Ископаемые брюхоногие моллюски были менее распространены в палеозойскую эпоху, чем двустворчатые моллюски . [29]
Большинство брюхоногих моллюсков палеозойской эры относятся к примитивным группам, некоторые из которых сохранились до сих пор. К каменноугольному периоду многие формы, наблюдаемые у современных брюхоногих моллюсков, могут быть сопоставлены с летописью окаменелостей, но, несмотря на это сходство во внешнем виде, большинство этих старых форм не имеют прямого отношения к живым формам. Именно в мезозойскую эру появились предки многих ныне живущих брюхоногих моллюсков. [29]
Одним из самых ранних известных наземных (наземных) брюхоногих моллюсков является Anthracopupa (= Maturipupa ), [30] который встречается в угольных мерах каменноугольного периода в Европе, но родственники современных наземных улиток до мелового периода редки . [29]
В породах мезозойской эры брюхоногие моллюски встречаются несколько чаще в виде ископаемых; их раковины часто хорошо сохраняются. Их окаменелости встречаются в древних пластах, отложенных как в пресноводной, так и в морской среде. « Мрамор Пурбека » юрского периода и « Мрамор Сассекса » раннего мелового периода, которые оба встречаются в южной Англии, представляют собой известняки , содержащие плотно упакованные останки прудовика Viviparus . [29]
Породы кайнозойской эры содержат очень большое количество окаменелостей брюхоногих моллюсков, многие из этих окаменелостей тесно связаны с современными живыми формами. Разнообразие брюхоногих моллюсков заметно увеличилось в начале этой эры вместе с разнообразием двустворчатых моллюсков . [29]
Некоторые следы, сохранившиеся в древних осадочных породах, как полагают, были оставлены брюхоногими моллюсками, ползавшими по мягкой грязи и песку. Хотя эти следы окаменелостей имеют спорное происхождение, некоторые из них действительно напоминают следы, оставленные современными брюхоногими моллюсками. [29]
Окаменелости брюхоногих моллюсков иногда можно спутать с аммонитами или другими головоногими моллюсками с панцирем . Примером тому является Беллерофонт из известняков каменноугольного периода Европы, раковина которого плоскоспирально извита и может быть принята за панцирь головоногих моллюсков. [ нужна цитата ]
Брюхоногие моллюски — одна из групп, фиксирующих изменения фауны, вызванные наступлением и отступлением ледниковых щитов в эпоху плейстоцена .
Кладограмма , показывающая филогенетические связи Gastropoda с примерами видов: [31]
Cocculiniformia , Neomphalina и Lower Heterobranchia не включены в приведенную кладограмму.
Современная классификация брюхоногих опирается на результаты филогеномного анализа. Консенсус еще не достигнут относительно взаимоотношений в самом основании древа жизни брюхоногих моллюсков, но в остальном основные группы известны с уверенностью. [32] [33] [34]



Со времен Дарвина биологическая систематика пыталась отразить филогению организмов, т. е. древо жизни . Классификации, используемые в таксономии, пытаются отразить точную взаимосвязь различных таксонов. Однако таксономия брюхоногих постоянно пересматривается, поэтому версии, представленные в различных текстах, могут существенно отличаться.
В старой классификации брюхоногих было четыре подкласса: [35]
Таксономия брюхоногих все еще находится на стадии пересмотра, и от старой таксономии отказываются все больше и больше, поскольку результаты исследований ДНК постепенно становятся все более ясными . Тем не менее, некоторые из старых терминов, таких как «заднежаберный» и «переднежаберный», до сих пор иногда используются в описательном смысле.
Новые открытия, основанные на секвенировании ДНК брюхоногих моллюсков, привели к некоторым революционным таксономическим открытиям. В случае брюхоногих таксономия сейчас постепенно переписывается, чтобы включить строго монофилетические группы (только одна линия брюхоногих моллюсков в каждой группе). Интеграция новых результатов в рабочую таксономию остается сложной задачей. От последовательных рангов внутри таксономии на уровне подкласса, надотряда, отряда и подотряда уже отказались как неработоспособных. В ближайшем будущем ожидается постоянный пересмотр более высоких таксономических уровней.
Конвергентная эволюция , которая, по-видимому, особенно часто встречается у брюхоногих моллюсков, может объяснить наблюдаемые различия между более старыми филогениями, основанными на морфологических данных, и более поздними исследованиями секвенирования генов.
В 2004 году Брайан Симисон и Дэвид Р. Линдберг показали возможное дифилетическое происхождение брюхоногих моллюсков на основе порядка митохондриальных генов и анализа аминокислотной последовательности полных генов. [36]
В 2005 году Филипп Буше и Жан-Пьер Рокруа внесли радикальные изменения в систематику , в результате чего была создана таксономия Буше и Рокруа , которая является шагом ближе к эволюционной истории этого типа . [3] [37] Система классификации Буше и Рокруа основана частично на старых системах классификации, а частично на новых кладистических исследованиях. В прошлом систематика брюхоногих во многом основывалась на фенетических морфологических признаках таксонов. Последние достижения в большей степени основаны на молекулярных характеристиках ДНК [38] и исследованиях РНК . Это сделало таксономические ранги и их иерархию спорными.
В 2017 году Буше, Рокруа и другие сотрудники опубликовали значительно обновленную версию таксономии 2005 года. [39] В таксономии Буше и др . авторы использовали нерангированные клады для таксонов выше ранга надсемейства (заменяя ранги подотряда, порядка, надотряда и подкласса), при этом используя традиционный подход Линнея для всех таксонов ниже ранга надсемейства. Во всех случаях, когда монофилия не проверялась или было известно, что она является парафилетической или полифилетической, использовался термин «группа» или «неформальная группа». Классификация семейств на подсемейства часто не совсем решена. [ нужна цитата ]
Многие виды брюхоногих моллюсков сталкиваются с угрозами разрушения среды обитания, загрязнения и изменения климата. Некоторые виды находятся под угрозой исчезновения или вымерли из-за этих факторов. Усилия по сохранению часто направлены на защиту среды их обитания, особенно в пресноводных и наземных экосистемах.
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ){kind=link}