
Паразитизм — это тесная связь между видами , при которой один организм, паразит, живет на другом организме, хозяине , или внутри него , причиняя ему определенный вред, и структурно адаптирован к такому образу жизни. [1] Энтомолог Э. О. Уилсон охарактеризовал паразитов как «хищников, которые поедают добычу группами менее одного». [2] Паразиты включают одноклеточных простейших, таких как возбудители малярии , сонной болезни и амебной дизентерии ; животных, таких как анкилостомы , вши , комары и летучие мыши-вампиры ; грибы, такие как опенок и возбудители стригущего лишая ; и растения, такие как омела , повилика и заразиха .
Существует шесть основных паразитарных стратегий эксплуатации животных-хозяев, а именно паразитарная кастрация , паразитизм, передающийся напрямую (при контакте), паразитизм, передающийся трофически (при поедании), паразитизм, передающийся через вектор , паразитоидизм и микрохищничество. Одна из основных осей классификации касается инвазивности: эндопаразит живет внутри тела хозяина; эктопаразит живет снаружи, на поверхности хозяина.
Как и хищничество, паразитизм является типом взаимодействия потребителя и ресурса [3], но в отличие от хищников паразиты, за исключением паразитоидов, намного меньше своих хозяев, не убивают их и часто живут в или на своих хозяевах в течение длительного периода. Паразиты животных высокоспециализированы , каждый вид паразита живет на одном данном виде животных и размножается с большей скоростью, чем их хозяева. Классическими примерами являются взаимодействия между позвоночными хозяевами и ленточными червями , сосальщиками , а также между видами Plasmodium , вызывающими малярию , и блохами .
Паразиты снижают приспособленность хозяина посредством общей или специализированной патологии , которая варьируется от паразитарной кастрации до модификации поведения хозяина . Паразиты повышают собственную приспособленность, эксплуатируя хозяев для получения ресурсов, необходимых для их выживания, в частности, питаясь ими и используя промежуточных (вторичных) хозяев для содействия их передаче от одного окончательного (первичного) хозяина к другому. Хотя паразитизм часто недвусмыслен, он является частью спектра взаимодействий между видами , переходящих через паразитоидизм в хищничество, через эволюцию в мутуализм , а у некоторых грибов переходя в сапрофитию .
Человеческие знания о паразитах, таких как круглые черви и ленточные черви, восходят к Древнему Египту , Греции и Риму . В ранние современные времена Антони ван Левенгук наблюдал Giardia lamblia с помощью своего микроскопа в 1681 году, в то время как Франческо Реди описал внутренних и внешних паразитов, включая овечью двуустку и клещей . Современная паразитология развилась в 19 веке. В человеческой культуре паразитизм имеет негативные коннотации. Они были использованы в сатирическом эффекте в поэме Джонатана Свифта 1733 года «О поэзии: Рапсодия», где поэты сравнивались с гиперпаразитарными «паразитами». В художественной литературе готический роман ужасов Брэма Стокера 1897 года «Дракула » и его многочисленные более поздние адаптации изображали кровососущего паразита. Фильм Ридли Скотта 1979 года «Чужой» был одним из многих произведений научной фантастики, в которых фигурировал паразитический инопланетный вид. [4]
Впервые использованное в английском языке в 1539 году, слово parasite происходит от средневекового французского parasite , от латинизированной формы parasitus , от древнегреческого παράσιτος [5] (parasitos) «тот, кто ест за столом другого», в свою очередь от παρά [6] (para) «рядом, около» и σῖτος (sitos) «пшеница, еда». [7] Родственный термин parasitism появляется в английском языке с 1611 года. [8]
Паразитизм — это разновидность симбиоза , тесного и постоянного долгосрочного биологического взаимодействия между паразитом и его хозяином. В отличие от сапротрофов , паразиты питаются живыми хозяевами, хотя некоторые паразитические грибы, например, могут продолжать питаться хозяевами, которых они убили. В отличие от комменсализма и мутуализма , паразитические отношения наносят вред хозяину, либо питаясь им, либо, как в случае кишечных паразитов, потребляя часть его пищи. Поскольку паразиты взаимодействуют с другими видами, они могут легко выступать в качестве переносчиков патогенов, вызывая заболевания . [9] [10] [11] Хищничество по определению не является симбиозом, поскольку взаимодействие кратковременно, но энтомолог Э. О. Уилсон охарактеризовал паразитов как «хищников, которые поедают добычу группами менее одного». [2]
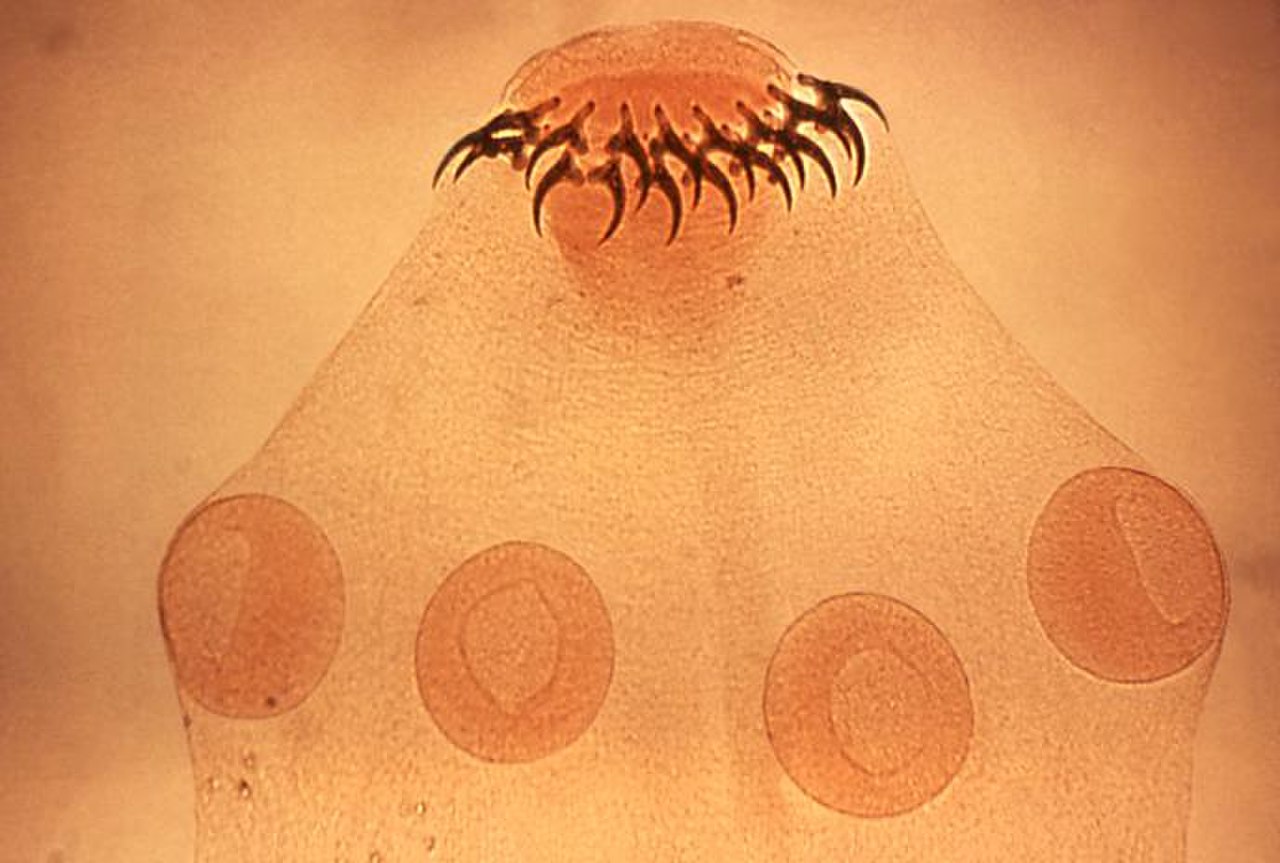
В этой области существует множество возможных стратегий. Таксономисты классифицируют паразитов по множеству перекрывающихся схем, основанных на их взаимодействии с хозяевами и на их жизненных циклах , которые иногда бывают очень сложными. Облигатный паразит полностью зависит от хозяина для завершения своего жизненного цикла, в то время как факультативный паразит этого не делает. Жизненные циклы паразитов, включающие только одного хозяина, называются «прямыми»; те, в которых есть окончательный хозяин (где паразит размножается половым путем) и по крайней мере один промежуточный хозяин, называются «косвенными». [12] [13] Эндопаразит живет внутри тела хозяина; эктопаразит живет снаружи, на поверхности хозяина. [14] Мезопаразиты, например, некоторые веслоногие рачки , проникают в отверстие в теле хозяина и частично остаются там. [15] Некоторые паразиты могут быть универсалами, питаясь широким спектром хозяев, но многие паразиты и большинство простейших и гельминтов , паразитирующих на животных, являются специалистами и крайне специфичны для хозяина. [14] Раннее базовое функциональное разделение паразитов различало микропаразитов и макропаразитов. Каждому из них была назначена математическая модель для анализа перемещений популяций групп хозяин-паразит. [16] Микроорганизмы и вирусы, которые могут размножаться и завершать свой жизненный цикл внутри хозяина, известны как микропаразиты. Макропаразиты — это многоклеточные организмы, которые размножаются и завершают свой жизненный цикл вне хозяина или на теле хозяина. [16] [17]
Большая часть размышлений о типах паразитизма была сосредоточена на наземных животных-паразитах животных, таких как гельминты. Те, кто находится в других средах и с другими хозяевами, часто имеют аналогичные стратегии. Например, курносый угорь, вероятно, является факультативным эндопаразитом (т. е. полупаразитом), который оппортунистически зарывается в больную и умирающую рыбу и поедает ее. [18] Растительноядные насекомые, такие как щитовки , тли и гусеницы , очень похожи на эктопаразитов, нападая на гораздо более крупные растения; они служат переносчиками бактерий, грибков и вирусов, вызывающих заболевания растений . Поскольку самки щитовок не могут двигаться, они являются облигатными паразитами, постоянно прикрепленными к своим хозяевам. [16]
Сенсорные сигналы, которые паразит использует для идентификации и приближения к потенциальному хозяину, известны как «сигналы хозяина». Такие сигналы могут включать, например, вибрацию, [19] выдыхаемый углекислый газ , запахи кожи, визуальные и тепловые сигнатуры и влажность. [20] Паразитические растения могут использовать, например, свет, физиохимию хозяина и летучие вещества для распознавания потенциальных хозяев. [21]
Существует шесть основных паразитарных стратегий , а именно паразитарная кастрация ; паразитизм, передающийся напрямую; паразитизм, передающийся трофически; паразитизм, передающийся вектором ; паразитоидизм ; и микрохищничество. Они применимы к паразитам, хозяевами которых являются как растения, так и животные. [16] [22] Эти стратегии представляют собой адаптивные пики ; возможны промежуточные стратегии, но организмы во многих различных группах последовательно сходились на этих шести, которые являются эволюционно стабильными. [22]
Перспективу эволюционных вариантов можно получить, рассмотрев четыре ключевых вопроса: влияние на приспособленность хозяев паразита; количество хозяев, которые у них есть на каждой стадии жизни; мешает ли хозяину размножаться; и зависит ли эффект от интенсивности (количества паразитов на хозяина). Из этого анализа вытекают основные эволюционные стратегии паразитизма, наряду с хищничеством. [23]

Паразитические кастраторы частично или полностью уничтожают способность своего хозяина к размножению, направляя энергию, которая пошла бы на размножение, на рост хозяина и паразита, иногда вызывая гигантизм у хозяина. Другие системы хозяина остаются нетронутыми, что позволяет ему выживать и поддерживать паразита. [22] [24] Паразитические ракообразные, такие как представители специализированного рода морских желудей Sacculina, в частности, наносят ущерб гонадам своих многочисленных видов [25] крабов -хозяев . В случае Sacculina яички более двух третей их хозяев-крабов дегенерируют в достаточной степени для того, чтобы у этих самцов-крабов развились вторичные половые признаки самок , такие как более широкие брюшки, меньшие клешни и придатки для захвата яиц. Различные виды гельминтов кастрируют своих хозяев (например, насекомых и улиток). Это может происходить напрямую, либо механически, питаясь их гонадами, либо выделяя химическое вещество, разрушающее репродуктивные клетки; или косвенно, будь то путем секреции гормона или путем отвода питательных веществ. Например, трематода Zoogonus lasius , чьи спороцисты лишены рта, химически кастрирует приливную морскую улитку Tritia obsoleta , развиваясь в ее гонаде и убивая ее репродуктивные клетки. [24] [26]

Паразиты, передающиеся напрямую, не требующие переносчика для достижения своих хозяев, включают таких паразитов наземных позвоночных, как вши и клещи; морских паразитов, таких как веслоногие рачки и циамидные амфиподы; моногенеи ; и многие виды нематод, грибов, простейших, бактерий и вирусов. Будь то эндопаразиты или эктопаразиты, у каждого есть один вид-хозяин. В пределах этого вида большинство особей свободны или почти свободны от паразитов, в то время как меньшинство переносит большое количество паразитов; это известно как агрегированное распределение . [22]

Трофически передаваемые паразиты передаются через поедание хозяином. К ним относятся трематоды (все, кроме шистосом ), цестоды , скребни , пентастомиды , многие круглые черви и многие простейшие, такие как токсоплазма . [22] У них сложные жизненные циклы, включающие хозяев двух или более видов. На ювенильных стадиях они заражают и часто инцистируют промежуточного хозяина. Когда промежуточное животное-хозяин съедается хищником, окончательным хозяином, паразит переживает процесс пищеварения и созревает во взрослую особь; некоторые живут как кишечные паразиты . Многие трофически передаваемые паразиты изменяют поведение своих промежуточных хозяев, увеличивая их шансы быть съеденными хищником. Как и в случае с напрямую передаваемыми паразитами, распределение трофически передаваемых паразитов среди особей хозяина является агрегированным. [22] Коинфекция несколькими паразитами является обычным явлением. [27] Аутоинфекция , при которой (в виде исключения) весь жизненный цикл паразита происходит в одном первичном хозяине, иногда может происходить у гельминтов, таких как Strongyloides stercoralis . [28]

Паразиты , передающиеся через переносчиков, полагаются на третью сторону, промежуточного хозяина, где паразит не размножается половым путем, [14] чтобы переносить их от одного окончательного хозяина к другому. [22] Эти паразиты представляют собой микроорганизмы, а именно простейшие , бактерии или вирусы , часто внутриклеточные патогены (возбудители болезней). [22] Их переносчиками в основном являются гематофагические членистоногие, такие как блохи, вши, клещи и комары. [22] [29] Например, олений клещ Ixodes scapularis выступает в качестве переносчика таких заболеваний, как болезнь Лайма , бабезиоз и анаплазмоз . [30] Простейшие эндопаразиты, такие как малярийные паразиты рода Plasmodium и паразиты сонной болезни рода Trypanosoma , имеют инфекционные стадии в крови хозяина, которые переносятся к новым хозяевам через укусы насекомых. [31]
Паразитоиды — это насекомые, которые рано или поздно убивают своих хозяев, что делает их отношения близкими к хищничеству. [32] Большинство паразитоидов — это паразитоидные осы или другие перепончатокрылые ; другие включают двукрылых, таких как мухи-фориды . Их можно разделить на две группы: идиобионты и койнобионты, различающиеся по своему отношению к хозяевам. [33]
Паразитоиды -идиобионты жалят свою часто крупную добычу при поимке, убивая ее на месте или немедленно парализуя. Затем обездвиженная добыча переносится в гнездо, иногда вместе с другой добычей, если она недостаточно велика, чтобы поддерживать паразитоида на протяжении всего его развития. Яйцо откладывается поверх добычи, а затем гнездо запечатывается. Паразитоид быстро развивается через стадии личинки и куколки, питаясь оставленными для него запасами. [33]
Паразитоиды койнобионтов , к которым относятся мухи и осы, откладывают яйца внутри молодых хозяев, обычно личинок. Им позволяют расти, поэтому хозяин и паразитоид развиваются вместе в течение длительного периода, заканчиваясь, когда паразитоиды вырастают во взрослых особей, оставляя добычу мертвой, съеденной изнутри. Некоторые койнобионты регулируют развитие своего хозяина, например, не давая ему окукливаться или заставляя его линять, когда паразитоид готов к линьке. Они могут делать это, вырабатывая гормоны, имитирующие гормоны линьки хозяина ( экдистероиды ), или регулируя эндокринную систему хозяина. [33]


Микрохищник атакует более одного хозяина, по крайней мере, на небольшую величину снижая приспособленность каждого хозяина, и контактирует с любым одним хозяином только периодически. Такое поведение делает микрохищников подходящими переносчиками, поскольку они могут передавать более мелких паразитов от одного хозяина к другому. [22] [34] [23] Большинство микрохищников являются гематофагами , питающимися кровью. К ним относятся кольчатые черви, такие как пиявки , ракообразные, такие как бранхиуры и изоподы гнатииды , различные двукрылые , такие как комары и мухи цеце , другие членистоногие, такие как блохи и клещи, позвоночные, такие как миноги , и млекопитающие, такие как летучие мыши-вампиры . [22]

Паразиты используют различные методы заражения животных-хозяев, включая физический контакт, фекально-оральный путь , свободноживущие инфекционные стадии и векторы, подходящие для их различных хозяев, жизненных циклов и экологических контекстов. [35] Примеры, иллюстрирующие некоторые из многих возможных комбинаций, приведены в таблице.
Среди многочисленных вариаций паразитических стратегий можно выделить гиперпаразитизм, [37] социальный паразитизм, [38] выводковый паразитизм, [39] клептопаразитизм, [40] половой паразитизм, [41] и адельфопаразитизм. [42]
Гиперпаразиты питаются другим паразитом, как показано на примере простейших, живущих в гельминтных паразитах, [37] или факультативных или облигатных паразитоидах, хозяевами которых являются либо обычные паразиты, либо паразитоиды. [22] [33] Уровни паразитизма за пределами вторичного также встречаются, особенно среди факультативных паразитоидов. В системах галлов дуба может быть до пяти уровней паразитизма. [43]
Гиперпаразиты могут контролировать популяции своих хозяев и используются для этой цели в сельском хозяйстве и в некоторой степени в медицине . Контролирующие эффекты можно увидеть в том, как вирус CHV1 помогает контролировать ущерб, который каштановая гниль , Cryphonectria parasitica , наносит американским каштанам , и в том, как бактериофаги могут ограничивать бактериальные инфекции. Вероятно, хотя и мало исследовано, что большинство патогенных микропаразитов имеют гиперпаразитов, которые могут оказаться широко полезными как в сельском хозяйстве, так и в медицине. [44]
Социальные паразиты используют в своих интересах межвидовые взаимодействия между членами эусоциальных животных, такими как муравьи , термиты и шмели . Примерами служат большая голубая бабочка Phengaris arion , личинки которой используют мимикрию муравьев для паразитирования на определенных муравьях, [38] Bombus bohemicus , шмель, который вторгается в ульи других пчел и берет на себя воспроизводство, пока их детеныши выращиваются рабочими-хозяевами, и Melipona scutellaris , эусоциальная пчела, чьи неплодные королевы избегают рабочих-убийц и вторгаются в другую колонию без королевы. [45] Крайний пример межвидового социального паразитизма обнаружен у муравья Tetramorium inquilinum , облигатного паразита, который живет исключительно на спинах других муравьев Tetramorium . [46] Механизм эволюции социального паразитизма был впервые предложен Карло Эмери в 1909 году. [47] Теперь известный как « правило Эмери », он гласит, что социальные паразиты, как правило, тесно связаны со своими хозяевами, часто принадлежа к одному роду. [48] [49] [50]
Внутривидовой социальный паразитизм происходит при паразитическом вскармливании, когда некоторые детеныши берут молоко у неродственных самок. У клиновидных капуцинов самки более высокого ранга иногда берут молоко у самок более низкого ранга без какой-либо взаимности. [51]
При гнездовом паразитизме хозяева выступают в роли родителей, поскольку они воспитывают детенышей как своих собственных. К гнездовым паразитам относятся птицы из разных семейств, такие как коровьи птицы , уточки , кукушки и черноголовые утки . Они не строят собственных гнезд, а оставляют свои яйца в гнездах других видов . Яйца некоторых гнездовых паразитов имитируют яйца своих хозяев, в то время как яйца некоторых коровьих птиц имеют прочную скорлупу, что затрудняет их уничтожение хозяевами путем прокалывания, оба механизма подразумевают отбор хозяевами паразитических яиц. [39] [52] [53] Взрослая самка европейской кукушки далее имитирует хищника, европейского перепелятника , давая ей время отложить свои яйца в гнезде хозяина незаметно. [54]
При клептопаразитизме (от греч. κλέπτης ( kleptēs ), «вор») паразиты крадут пищу, собранную хозяином. Паразитизм часто происходит на близких родственниках, будь то в пределах одного вида или между видами в одном роде или семействе. Например, многочисленные линии пчел-кукушек откладывают яйца в ячейки гнезд других пчел в том же семействе. [40] Клептопаразитизм в целом встречается редко, но заметен у птиц; некоторые, такие как поморники, специализируются на пиратстве пищи у других морских птиц, неустанно преследуя их, пока они не извергнут свою добычу. [55]
Уникальный подход наблюдается у некоторых видов удильщиков , таких как Ceratias holboelli , где самцы сводятся к крошечным половым паразитам , полностью зависящим от самок своего вида для выживания, постоянно прикрепленным ниже тела самки и неспособным позаботиться о себе. Самка кормит самца и защищает его от хищников, в то время как самец ничего не дает взамен, кроме спермы, которая нужна самке для производства следующего поколения. [41]
Адельфопаразитизм (от греч. ἀδελφός ( adelphós ), брат [56] ), также известный как сиблинг-паразитизм, происходит, когда вид хозяина тесно связан с паразитом, часто в том же семействе или роде. [42] У паразитоида цитрусовой мошки Encarsia perplexa неспаренные самки могут откладывать гаплоидные яйца в полностью развитых личинках своего собственного вида, производя мужское потомство, [57] в то время как морской червь Bonellia viridis имеет схожую репродуктивную стратегию, хотя личинки являются планктонными. [58]
Приведены примеры основных вариантов стратегий.
Паразитизм имеет чрезвычайно широкий таксономический диапазон, включающий животных, растения, грибы, простейших, бактерии и вирусы. [59]
Паразитизм широко распространен в животном мире [63] и сотни раз развивался независимо от свободноживущих форм. [22] Многие типы гельминтов, включая сосальщиков и цестод, имеют полные жизненные циклы, включающие двух или более хозяев. Безусловно, самая большая группа — это паразитоидные осы среди перепончатокрылых. [22] Типы и классы с наибольшим количеством паразитических видов перечислены в таблице. Числа представляют собой консервативные минимальные оценки. Столбцы для эндо- и эктопаразитизма относятся к окончательному хозяину, как указано в столбцах позвоночных и беспозвоночных. [60]

Полупаразит или частичный паразит, такой как омела, получает часть своих питательных веществ из другого живого растения, тогда как голопаразит, такой как повилика, получает все свои питательные вещества из другого растения. [64] Паразитические растения составляют около одного процента покрытосеменных растений и встречаются почти в каждом биоме мира. [65] [66] [67] Все эти растения имеют модифицированные корни, гаустории , которые проникают в растения-хозяева, соединяя их с проводящей системой — либо ксилемой , либо к флоэме , либо к обеим. Это дает им возможность извлекать воду и питательные вещества из хозяина. Паразитическое растение классифицируется в зависимости от того, где оно прикрепляется к хозяину, либо к стеблю, либо к корню, и от количества необходимых ему питательных веществ. Поскольку голопаразиты не имеют хлорофилла и, следовательно, не могут производить для себя пищу путем фотосинтеза , они всегда являются облигатными паразитами, получающими всю свою пищу от своих хозяев. [66] Некоторые паразитические растения могут определять местонахождение своих растений- хозяев , обнаруживая химические вещества в воздухе или почве, выделяемые побегами или корнями хозяина , соответственно. Известно около 4500 видов паразитических растений примерно в 20 семействах цветковых растений . [68] [66]
Виды семейства Orobanchaceae (заразихи) являются одними из самых экономически разрушительных из всех растений. Виды Striga (ведьминых сорняков) оцениваются в миллиарды долларов в год из-за потери урожая, заражая более 50 миллионов гектаров возделываемых земель только в странах Африки к югу от Сахары. Striga заражает как травы, так и зерновые, включая кукурузу , рис и сорго , которые являются одними из важнейших продовольственных культур в мире. Orobanche также угрожает широкому спектру других важных культур, включая горох , нут , томаты , морковь и разновидности капусты . Потеря урожая от Orobanche может быть полной; несмотря на обширные исследования, ни один метод контроля не был полностью успешным. [69]
Многие растения и грибы обмениваются углеродом и питательными веществами в мутуалистических микоризных отношениях. Однако около 400 видов микогетеротрофных растений, в основном в тропиках, эффективно обманывают , забирая углерод у гриба, а не обменивая его на минералы. У них сильно редуцированные корни, так как им не нужно поглощать воду из почвы; их стебли тонкие с небольшим количеством сосудистых пучков , а их листья редуцированы до мелких чешуек, так как они не фотосинтезируют. Их семена очень мелкие и многочисленные, поэтому они, по-видимому, полагаются на заражение подходящим грибом вскоре после прорастания. [70]

Паразитические грибы получают часть или все необходимые им питательные вещества из растений, других грибов или животных.
Фитопатогенные грибы подразделяются на три категории в зависимости от способа питания: биотрофы, гемибиотрофы и некротрофы. Биотрофные грибы получают питательные вещества из живых растительных клеток, и в ходе заражения они колонизируют свое растение-хозяина таким образом, чтобы поддерживать его живым в течение максимально длительного времени. [71] Одним из известных примеров биотрофного патогена является Ustilago maydis , возбудитель головневой болезни кукурузы. Некротрофные патогены, с другой стороны, убивают клетки-хозяева и питаются сапрофитно , примером являются колонизирующие корни медовые грибы рода Armillaria . [72] Гемибиотрофные патогены начинают колонизацию своих хозяев как биотрофы, а затем убивают клетки-хозяева и питаются как некротрофы, явление, называемое переключением биотрофия-некротрофия. [73]
Патогенные грибы являются хорошо известными возбудителями заболеваний как у животных, так и у людей. По оценкам, грибковые инфекции ( микозы ) убивают 1,6 миллиона человек каждый год. [74] Одним из примеров сильнодействующих грибковых патогенов животных являются Microsporidia — облигатные внутриклеточные паразитические грибы, которые в основном поражают насекомых, но могут также поражать позвоночных, включая людей, вызывая кишечную инфекцию микроспоридиоз . [75]

Простейшие, такие как Plasmodium , Trypanosoma и Entamoeba [76], являются эндопаразитарными. Они вызывают серьезные заболевания у позвоночных, включая людей, — в этих примерах малярию, сонную болезнь и амебную дизентерию — и имеют сложные жизненные циклы. [31]
Многие бактерии являются паразитическими, хотя их чаще считают патогенами, вызывающими заболевания. [77] Паразитические бактерии чрезвычайно разнообразны и заражают своих хозяев различными путями. Приведем несколько примеров: Bacillus anthracis , возбудитель сибирской язвы , распространяется при контакте с инфицированными домашними животными ; ее споры , которые могут годами выживать вне организма, могут попасть в организм хозяина через ссадины или могут быть вдыхаемы. Borrelia , возбудитель болезни Лайма и возвратного тифа , передается переносчиками, клещами рода Ixodes , из резервуаров болезней у животных, таких как олени . Campylobacter jejuni , возбудитель гастроэнтерита , распространяется фекально-оральным путем от животных или при употреблении в пищу недостаточно приготовленной птицы или через загрязненную воду. Haemophilus influenzae , возбудитель бактериального менингита и инфекций дыхательных путей, таких как грипп и бронхит , передается воздушно-капельным путем. Treponema pallidum , возбудитель сифилиса , распространяется половым путем . [ 78]

Вирусы являются облигатными внутриклеточными паразитами, характеризующимися крайне ограниченной биологической функцией, вплоть до того, что, хотя они, очевидно, способны заражать все другие организмы от бактерий и архей до животных, растений и грибов, неясно, можно ли их самих описать как живые. Они могут быть как РНК- , так и ДНК-вирусами, состоящими из одинарной или двойной цепи генетического материала ( РНК или ДНК соответственно), покрытыми белковой оболочкой, а иногда и липидной оболочкой. Таким образом, у них отсутствуют все обычные механизмы клетки, такие как ферменты , и они полностью полагаются на способность клетки-хозяина реплицировать ДНК и синтезировать белки. Большинство вирусов являются бактериофагами , заражающими бактерии. [79] [80] [81] [82]

Паразитизм является важным аспектом эволюционной экологии; например, почти все свободноживущие животные являются хозяевами по крайней мере одного вида паразита. Позвоночные, наиболее изученная группа, являются хозяевами от 75 000 до 300 000 видов гельминтов и неисчислимого количества паразитических микроорганизмов. В среднем, вид млекопитающих является хозяином четырех видов нематод, двух видов трематод и двух видов цестод. [83] У людей есть 342 вида паразитических гельминтов и 70 видов простейших паразитов. [84] Около трех четвертей связей в пищевых сетях включают паразита, важного для регулирования численности хозяев. Возможно, 40 процентов описанных видов являются паразитами. [83]
Паразитизм трудно продемонстрировать на основе ископаемых остатков , но отверстия в челюстях нескольких образцов тираннозавра могли быть вызваны паразитами, подобными трихомонадам . [85] Saurophthirus , раннемеловая блоха , паразитировала на птерозаврах . [86] [87] Яйца, принадлежавшие нематодам , и, вероятно, цисты простейших были обнаружены в позднетриасовом копролите фитозавра . Эта редкая находка в Таиланде раскрывает больше информации об экологии доисторических паразитов. [ 88]
Поскольку хозяева и паразиты развиваются вместе, их отношения часто меняются. Когда паразит находится в единственных отношениях с хозяином, отбор делает отношения более благоприятными, даже мутуалистическими, поскольку паразит может размножаться дольше, если его хозяин живет дольше. [89] Но когда паразиты конкурируют, отбор благоприятствует паразиту, который размножается быстрее, что приводит к повышению вирулентности. Таким образом, существуют различные возможности в коэволюции хозяина и паразита . [90]
Эволюционная эпидемиология анализирует, как паразиты распространяются и развиваются, тогда как дарвиновская медицина применяет схожее эволюционное мышление к непаразитарным заболеваниям, таким как рак и аутоиммунные состояния . [91]

Долгосрочные партнерства могут привести к относительно стабильным отношениям, имеющим тенденцию к комменсализму или мутуализму , поскольку при прочих равных условиях в эволюционном интересе паразита, чтобы его хозяин процветал. Паразит может эволюционировать, чтобы стать менее вредным для своего хозяина, или хозяин может эволюционировать, чтобы справиться с неизбежным присутствием паразита — до такой степени, что отсутствие паразита наносит вред хозяину. Например, хотя животные, паразитирующие на червях , часто явно страдают, такие инфекции могут также снижать распространенность и последствия аутоиммунных расстройств у животных-хозяев, включая людей. [89] В более экстремальном примере некоторые нематоды не могут размножаться или даже выживать без заражения бактериями Wolbachia . [92]
Линн Маргулис и другие утверждали, следуя книге Питера Кропоткина 1902 года « Взаимопомощь: фактор эволюции» , что естественный отбор меняет отношения от паразитизма к мутуализму, когда ресурсы ограничены. Этот процесс мог быть вовлечен в симбиогенез , который сформировал эукариот из внутриклеточных отношений между археями и бактериями, хотя последовательность событий остается в значительной степени неопределенной. [93] [94]
Можно ожидать, что конкуренция между паразитами будет способствовать более быстрому размножению и, следовательно, более опасным паразитам путем естественного отбора . [90] [95]
.jpg/1280px-Pink_Flamingos_with_Duck_-_Camargue,_France_-_April_2007_(cropped).jpg)
Среди конкурирующих паразитических бактерий, убивающих насекомых, родов Photorhabdus и Xenorhabdus вирулентность зависела от относительной силы антимикробных токсинов ( бактериоцинов ), вырабатываемых двумя вовлеченными штаммами. Когда только одна бактерия могла убить другую, другой штамм исключался из конкуренции. Но когда гусеницы были инфицированы бактериями, обе из которых имели токсины, способные убить другой штамм, ни один из штаммов не исключался, и их вирулентность была ниже, чем когда насекомое было инфицировано одним штаммом. [90]
Паразит иногда претерпевает совместное видообразование со своим хозяином, что приводит к закономерности, описанной в правиле Фаренхольца , согласно которой филогении хозяина и паразита начинают отражать друг друга. [96]
Примером может служить обезьяний пенистый вирус (SFV) и его хозяева-приматы. Было обнаружено, что филогении полимеразы SFV и субъединицы II митохондриальной цитохром с-оксидазы африканских и азиатских приматов тесно совпадают по порядку ветвления и времени расхождения, что подразумевает, что обезьяньи пенистые вирусы ковидировались с приматами Старого Света по крайней мере 30 миллионов лет. [97]
Предположение об общей эволюционной истории между паразитами и хозяевами может помочь выяснить, как связаны таксоны хозяев. Например, был спор о том, являются ли фламинго более тесно связанными с аистами или утками . Тот факт, что фламинго разделяют паразитов с утками и гусями, изначально принимался как доказательство того, что эти группы были более тесно связаны друг с другом, чем любая из них с аистами. Однако эволюционные события, такие как дупликация или вымирание видов паразитов (без схожих событий в филогении хозяина) часто разрушают сходство между филогениями хозяина и паразита. В случае фламинго, у них есть вши, похожие на вши поганок . У фламинго и поганок действительно есть общий предок, что подразумевает совместное видообразование птиц и вшей в этих группах. Затем вши фламинго сменили хозяев на уток, создав ситуацию, которая сбила с толку биологов. [98]
.jpg/1280px-Toxoplasma_gondii_(2).jpg)
Паразиты заражают симпатрических хозяев (находящихся в пределах их географической области) более эффективно, как было показано на примере дигенетических трематод, заражающих озерных улиток. [99] Это соответствует гипотезе Красной Королевы , которая гласит, что взаимодействие между видами приводит к постоянному естественному отбору для коадаптации. Паразиты отслеживают фенотипы локально распространенных хозяев, поэтому паразиты менее заразны для аллопатрических хозяев, которые находятся в других географических регионах. [99]
Некоторые паразиты изменяют поведение хозяина , чтобы увеличить свою передачу между хозяевами, часто в отношении хищника и добычи ( паразит усиливает трофическую передачу ). Например, в прибрежном солончаке Калифорнии трематода Euhaplorchis californiensis снижает способность своего хозяина -киллифа избегать хищников. [100] Этот паразит созревает в белых цаплях , которые с большей вероятностью питаются инфицированными киллифами, чем незараженной рыбой. Другим примером является простейшее Toxoplasma gondii , паразит, который созревает в кошках , но может переноситься многими другими млекопитающими . Неинфицированные крысы избегают запахов кошек, но крысы, инфицированные T. gondii, привлекаются этим запахом, что может увеличить передачу хозяевам-кошкам. [101] Малярийный паразит изменяет запах кожи своих хозяев-людей, увеличивая их привлекательность для комаров и, следовательно, повышая вероятность передачи паразита. [36] Паук Cyclosa argenteoalba часто имеет прикрепленных к ним личинок паразитоидных ос, которые изменяют их поведение при строительстве паутины. Вместо того, чтобы производить свои обычные липкие спиралевидные паутины, они создавали упрощенные паутины, когда паразиты были прикреплены. Это измененное поведение длилось дольше и было тем более заметным, чем дольше паразиты оставались на пауках. [102]

Паразиты могут эксплуатировать своих хозяев для выполнения ряда функций, которые им в противном случае пришлось бы выполнять самим. Паразиты, которые теряют эти функции, затем получают селективное преимущество, поскольку они могут перенаправлять ресурсы на воспроизводство. Многие эктопаразиты насекомых, включая клопов , клопов-летучих мышей , вшей и блох, утратили способность летать , полагаясь вместо этого на своих хозяев для транспортировки. [103] Потеря признаков в целом широко распространена среди паразитов. [104] Крайним примером является миксоспоридий Henneguya zschokkei , эктопаразит рыб и единственное животное, которое, как известно, утратило способность к аэробному дыханию: в его клетках отсутствуют митохондрии . [105]
Хозяева выработали множество защитных мер против своих паразитов, включая физические барьеры, такие как кожа позвоночных [106] , иммунная система млекопитающих [107], насекомые, активно удаляющие паразитов [108] , и защитные химические вещества в растениях [109] .
Эволюционный биолог У. Д. Гамильтон предположил, что половое размножение могло развиться, чтобы помочь победить множественных паразитов, обеспечивая генетическую рекомбинацию , перетасовку генов для создания различных комбинаций. Гамильтон показал с помощью математического моделирования, что половое размножение будет эволюционно стабильным в различных ситуациях, и что предсказания теории соответствуют фактической экологии полового размножения. [110] [111] Однако может существовать компромисс между иммунокомпетентностью и размножением вторичных половых признаков самцов позвоночных хозяев , таких как оперение павлинов и гривы львов . Это происходит потому, что мужской гормон тестостерон стимулирует рост вторичных половых признаков, благоприятствуя таким самцам в половом отборе , ценой снижения их иммунной защиты. [112]
.jpg/1280px-Short_Horned_Lizard_(4457945238).jpg)
Физический барьер жесткой и часто сухой и водонепроницаемой кожи рептилий, птиц и млекопитающих удерживает вторгающиеся микроорганизмы от проникновения в организм. Человеческая кожа также выделяет кожное сало , которое токсично для большинства микроорганизмов. [106] С другой стороны, более крупные паразиты, такие как трематоды, обнаруживают химические вещества, вырабатываемые кожей, чтобы обнаружить своих хозяев, когда они попадают в воду. Слюна и слезы позвоночных содержат лизоцим , фермент, который разрушает клеточные стенки вторгающихся бактерий. [106] Если организм проходит через рот, следующей линией обороны становится желудок с его соляной кислотой , токсичной для большинства микроорганизмов. [106] Некоторые кишечные паразиты имеют толстую, жесткую внешнюю оболочку, которая переваривается медленно или вообще не переваривается, позволяя паразиту проходить через желудок живым, после чего они попадают в кишечник и начинают следующую стадию своей жизни. Попав внутрь организма, паразиты должны преодолеть сывороточные белки иммунной системы и рецепторы распознавания образов , внутриклеточные и клеточные, которые активируют лимфоциты адаптивной иммунной системы, такие как Т-клетки и продуцирующие антитела В-клетки . У них есть рецепторы, которые распознают паразитов. [107]

Насекомые часто адаптируют свои гнезда, чтобы уменьшить паразитизм. Например, одна из основных причин, по которой оса Polistes canadensis гнездится в нескольких сотах , а не строит один гребень, как большинство других представителей ее рода, заключается в том, чтобы избежать заражения молью тинеид . Моль тинеид откладывает яйца в гнездах ос, а затем из этих яиц вылупляются личинки, которые могут рыть норы из ячейки в ячейку и охотиться на куколок ос. Взрослые осы пытаются удалить и убить яйца и личинки моли, пережевывая края ячеек, покрывая ячейки оральным секретом, который придает гнезду темно-коричневый вид. [108]
Растения реагируют на атаку паразита серией химических защит, таких как полифенолоксидаза , под контролем сигнальных путей жасмоновой кислоты, нечувствительной (JA) и салициловой кислоты (SA). [109] [113] Различные биохимические пути активируются различными атаками, и эти два пути могут взаимодействовать положительно или отрицательно. В целом, растения могут инициировать либо специфический, либо неспецифический ответ. [114] [113] Специфические ответы включают распознавание паразита клеточными рецепторами растения, что приводит к сильному, но локализованному ответу: защитные химикаты вырабатываются вокруг области, где был обнаружен паразит, блокируя его распространение и избегая напрасной траты защитного производства там, где это не нужно. [114] Неспецифические защитные ответы являются системными, что означает, что ответы не ограничиваются областью растения, а распространяются по всему растению, что делает их затратными с точки зрения энергии. Они эффективны против широкого спектра паразитов. [114] При повреждении, например, гусеницами чешуекрылых , листья растений, включая кукурузу и хлопок, выделяют повышенное количество летучих химических веществ, таких как терпены , которые сигнализируют о том, что они подвергаются нападению; одним из эффектов этого является привлечение паразитоидных ос, которые, в свою очередь, нападают на гусениц. [115]

Паразитизм и эволюция паразитов до двадцать первого века изучались паразитологами , в науке, в которой доминировала медицина, а не экологи или эволюционные биологи . Несмотря на то, что взаимодействия паразита и хозяина были явно экологическими и важными в эволюции, история паразитологии вызвала то, что эволюционный эколог Роберт Пулен назвал «захватом паразитизма паразитологами», что заставило экологов игнорировать эту область. По его мнению, это было «досадно», поскольку паразиты являются «вездесущими агентами естественного отбора» и значительными силами в эволюции и экологии. [116] По его мнению, давний раскол между науками ограничивал обмен идеями с отдельными конференциями и отдельными журналами. Технические языки экологии и паразитологии иногда включали разные значения для одних и тех же слов. Были и философские различия: Пулен отмечает, что под влиянием медицины «многие паразитологи признали, что эволюция привела к снижению вирулентности паразитов, тогда как современная эволюционная теория предсказала бы более широкий диапазон результатов» [116] .
Их сложные взаимоотношения затрудняют размещение паразитов в пищевых сетях: трематода с несколькими хозяевами для различных стадий жизненного цикла будет занимать много позиций в пищевой сети одновременно и будет создавать петли потока энергии, запутывая анализ. Кроме того, поскольку почти у каждого животного есть (множественные) паразиты, паразиты будут занимать верхние уровни каждой пищевой сети. [84]
Паразиты могут играть роль в распространении неместных видов. Например, инвазивные зеленые крабы минимально подвержены влиянию местных трематод на восточном побережье Атлантики. Это помогает им вытеснять местных крабов, таких как атлантические скальные и крабы-джоны . [117]
Экологическая паразитология может быть важна для попыток контроля, как во время кампании по искоренению ришты . Несмотря на то, что паразит был искоренен во всех странах, кроме четырех, червь начал использовать лягушек в качестве промежуточного хозяина, прежде чем заражать собак, что сделало контроль более сложным, чем если бы взаимосвязи были лучше поняты. [118]
Хотя паразиты широко считаются вредными, искоренение всех паразитов не принесет пользы. Паразиты составляют по крайней мере половину разнообразия жизни; они выполняют важные экологические роли; и без паразитов организмы могли бы стремиться к бесполому размножению, уменьшая разнообразие признаков, вызванных половым размножением. [119] Паразиты предоставляют возможность для передачи генетического материала между видами, способствуя эволюционным изменениям. [120] Многим паразитам требуется несколько хозяев разных видов для завершения их жизненных циклов и они полагаются на хищник-жертва или другие стабильные экологические взаимодействия, чтобы перейти от одного хозяина к другому. Таким образом, присутствие паразитов указывает на то, что экосистема здорова. [121]
Эктопаразит, калифорнийская кондоровая вошь, Colpocephalum californici , стал широко известной проблемой охраны природы. В Соединенных Штатах была запущена крупная и очень дорогостоящая программа разведения в неволе, чтобы спасти калифорнийского кондора . Он был хозяином воши, которая жила только за счет него. Все найденные вши были «намеренно убиты» во время программы, чтобы поддерживать кондоров в наилучшем возможном здоровье. Результатом стало то, что один вид, кондор, был спасен и возвращен в дикую природу, в то время как другой вид, паразит, вымер. [122]
Хотя паразиты часто опускаются в описаниях пищевых сетей , они обычно занимают верхнюю позицию. Паразиты могут функционировать как ключевые виды , снижая доминирование превосходящих конкурентов и позволяя конкурирующим видам сосуществовать. [84] [123] [124]

Один вид паразита обычно имеет агрегированное распределение среди животных-хозяев, что означает, что большинство хозяев переносят мало паразитов, в то время как несколько хозяев переносят подавляющее большинство особей паразитов. Это создает значительные проблемы для студентов, изучающих экологию паразитов, поскольку делает параметрическую статистику , обычно используемую биологами, недействительной. Несколько авторов рекомендуют логарифмическое преобразование данных перед применением параметрического теста или использование непараметрической статистики , но это может привести к дополнительным проблемам, поэтому количественная паразитология основана на более продвинутых биостатистических методах. [125]
Паразиты человека , включая круглых червей, ришту , острицы и ленточные черви, упоминаются в египетских папирусных записях с 3000 г. до н. э.; в папирусе Эберса описывается анкилостома . В Древней Греции паразиты, включая пузырчатого червя , описаны в Корпусе Гиппократа , в то время как комический драматург Аристофан называл ленточных червей «градинами». Римские врачи Цельс и Гален документировали круглых червей Ascaris lumbricoides и Enterobius vermicularis . [126]

В своем «Каноне врачебной науки» , завершенном в 1025 году, персидский врач Авиценна описал паразитов человека и животных, включая круглых червей, остриц, ришту и ленточных червей. [126]
В своей книге 1397 года «Traité de l'état, science et pratique de l'art de la Bergerie» («Очерк состояния, науки и практики искусства пастухов») Жан де Бри впервые описал эндопаразита трематоды — печеночного сосальщика овец Fasciola hepatica . [127] [128]
В ранний современный период в книге Франческо Реди 1668 года « Опыты зарождения насекомых » ( Esperienze Intorno alla Generazione degl'Insetti ) подробно описаны экто- и эндопаразиты, в частности клещи , личинки носовых мух оленей и печеночная двуустка овец . [129] Реди отметил, что паразиты развиваются из яиц, что противоречит теории спонтанного зарождения . [130] В своей книге 1684 года «Наблюдения за живыми животными, обнаруженные у живых животных» ( Osservazioni intorno agli animali viventi che si trovano negli animali viventi ) Реди описал и проиллюстрировал более 100 паразитов, включая большого круглого червя у людей, вызывающего аскаридоз . [129] Реди был первым, кто назвал цисты Echinococcus granulosus, обнаруженные у собак и овец, паразитическими; столетие спустя, в 1760 году, Питер Саймон Паллас правильно предположил, что это личинки ленточных червей. [126]
В 1681 году Антони ван Левенгук наблюдал и проиллюстрировал простейшего паразита Giardia lamblia и связал его со «своим жидким стулом». Это был первый простейший паразит человека, которого можно было увидеть под микроскопом. [126] Несколько лет спустя, в 1687 году, итальянские биологи Джованни Козимо Бономо и Диачинто Честони описали чесотку как вызываемую паразитическим клещом Sarcoptes scabiei , отметив ее как первое заболевание человека с известным микроскопическим возбудителем. [131]

Современная паразитология развивалась в 19 веке благодаря точным наблюдениям и экспериментам многих исследователей и врачей; [127] этот термин был впервые использован в 1870 году. [132] В 1828 году Джеймс Эннерсли описал амебиаз , протозойные инфекции кишечника и печени, хотя возбудитель, Entamoeba histolytica , был открыт только в 1873 году Фридрихом Лёшем. Джеймс Пэджет открыл кишечную нематоду Trichinella spiralis у людей в 1835 году. Джеймс Макконнелл описал человеческую печеночную двуустку, Clonorchis sinensis , в 1875 году. [126] Алджернон Томас и Рудольф Лейкарт независимо друг от друга сделали первое открытие жизненного цикла трематоды, овечьей печеночной двуустки, экспериментально в 1881–1883 годах. [127] В 1877 году Патрик Мэнсон открыл жизненный цикл филяриевых червей , которые вызывают слоновость, передаваемую комарами. Мэнсон также предсказал, что малярийный паразит, Plasmodium , имеет переносчика-комара, и убедил Рональда Росса провести исследование. Росс подтвердил, что предсказание было верным в 1897–1898 годах. В то же время Джованни Баттиста Грасси и другие описали стадии жизненного цикла малярийного паразита у комаров Anopheles . Росс был спорно награжден Нобелевской премией 1902 года за свою работу, в то время как Грасси не был. [126] В 1903 году Дэвид Брюс идентифицировал простейшего паразита и переносчика африканского трипаносомоза — муху цеце . [133]
Учитывая важность малярии, которой ежегодно заражаются около 220 миллионов человек, было предпринято много попыток прервать ее передачу. Были испробованы различные методы профилактики малярии , включая использование противомалярийных препаратов для уничтожения паразитов в крови, уничтожение ее переносчиков-комаров с помощью хлорорганических и других инсектицидов , а также разработка вакцины против малярии . Все они оказались проблематичными, с лекарственной устойчивостью , устойчивостью к инсектицидам среди комаров и повторной неэффективностью вакцин по мере мутации паразита. [134] Первой и по состоянию на 2015 год единственной лицензированной вакциной от любого паразитарного заболевания человека является RTS,S для малярии Plasmodium falciparum . [135]

Несколько групп паразитов, включая микробных патогенов и паразитоидных ос, использовались в качестве агентов биологической борьбы в сельском хозяйстве и садоводстве . [137] [138]
Пулен отмечает, что широко распространенное профилактическое использование антигельминтных препаратов у домашних овец и крупного рогатого скота представляет собой всемирный неконтролируемый эксперимент по эволюции жизненного цикла их паразитов. Результаты зависят от того, уменьшают ли препараты вероятность достижения личинки гельминта взрослой особи. Если это так, то можно ожидать, что естественный отбор будет благоприятствовать производству яиц в более раннем возрасте. Если, с другой стороны, препараты в основном воздействуют на взрослых паразитических червей , отбор может вызвать задержку созревания и повышенную вирулентность . Такие изменения, по-видимому, происходят: нематода Teladorsagia circumcincta меняет размер своей взрослой особи и скорость размножения в ответ на препараты. [139]

В классическую эпоху понятие паразита не было строго уничижительным: паразит был принятой ролью в римском обществе , в которой человек мог жить за счет гостеприимства других в обмен на «лесть, простые услуги и готовность терпеть унижения». [140] [141]
Паразитизм имеет уничижительный смысл в народном употреблении. По словам иммунолога Джона Плейфэра, [142]
В повседневной речи термин «паразит» нагружен уничижительным значением. Паразит — это нахлебник, ленивый наживальщик, обуздатель общества. [142]
Сатирик - священник Джонатан Свифт намекает на гиперпаразитизм в своей поэме 1733 года «О поэзии: Рапсодия», сравнивая поэтов с «паразитами», которые «дразнят и щипают своих врагов»: [143]
Паразиты только дразнят и щиплют
Своих врагов, превосходящих их на дюйм.
Так натуралисты замечают, что у блохи
Есть меньшие блохи, которые на нее охотятся;
А у этих есть меньшие блохи, чтобы кусать их.
И так продолжается до бесконечности .
Так каждый поэт, в своем роде,
Укушен тем, кто идет сзади:
Исследование 2022 года изучило названия около 3000 видов паразитов, открытых за предыдущие два десятилетия. Из названных в честь ученых, более 80% были названы в честь мужчин, тогда как около трети авторов статей о паразитах были женщинами. Исследование показало, что процент видов паразитов, названных в честь родственников или друзей автора, резко возрос за тот же период. [144]

В готическом романе ужасов Брэма Стокера 1897 года «Дракула » и его многочисленных экранизациях одноименный граф Дракула — кровососущий паразит (вампир). Критик Лора Отис утверждает, что как «вор, соблазнитель, творец и подражатель, Дракула — высший паразит. Весь смысл вампиризма — сосать чужую кровь, жить за счет других людей». [145]
Отвратительные и ужасающие паразитические инопланетные виды широко распространены в научной фантастике , [146] [147] как, например, в фильме Ридли Скотта 1979 года «Чужой» . [148] [149] В одной из сцен ксеноморф вырывается из груди мертвеца, и кровь под высоким давлением, которому способствуют взрывные устройства, вырывается наружу . Для усиления эффекта шока использовались органы животных . Сцена была снята одним дублем, и испуганная реакция актеров была подлинной. [4] [150]
Паразиты, если говорить кратко, это хищники, которые едят добычу группами менее одного экземпляра. Терпимые паразиты — это те, которые эволюционировали, чтобы обеспечить собственное выживание и воспроизводство, но в то же время с минимальными страданиями и затратами для хозяина.
Паразитизм — это форма симбиоза, тесная связь между двумя разными видами. Между хозяином и паразитом существует биохимическое взаимодействие; то есть они узнают друг друга, в конечном счете на молекулярном уровне, и ткани хозяина стимулируются к определенной реакции. Это объясняет, почему паразитизм может привести к болезни, но не всегда.
, травоядность и паразитизм существуют в континууме тяжести с точки зрения степени, в которой они отрицательно влияют на приспособленность организма. ... В большинстве ситуаций паразиты не убивают своих хозяев. Исключение, однако, составляют паразитоиды, которые стирают грань между паразитизмом и хищничеством.
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка )XIX век можно считать временем зарождения современной паразитологии.
естественно, сосредотачивается на отношениях с наибольшей зависимостью, на тех, в которых клиент получает пищу от своего покровителя, и для этого заранее сконструированная персона паразита оказалась чрезвычайно полезной.
![Идиобионтные паразитоидные осы немедленно парализуют своих хозяев, чтобы их личинки (Pimplinae, на фото) могли их съесть.[22]](http://upload.wikimedia.org/wikipedia/commons/thumb/6/65/Live_Tetragnatha_montana_(RMNH.ARA.14127)_parasitized_by_Acrodactyla_quadrisculpta_larva_(RMNH.INS.593867)_-_BDJ.1.e992.jpg/1280px-Live_Tetragnatha_montana_(RMNH.ARA.14127)_parasitized_by_Acrodactyla_quadrisculpta_larva_(RMNH.INS.593867)_-_BDJ.1.e992.jpg)




.jpg/1280px-Great_Skua_(cropped).jpg)
.jpg/1280px-%D0%A1%D0%B5%D0%B2%D0%B5%D1%80%D0%BD%D0%B0%D1%8F_%D1%86%D0%B5%D1%80%D0%B0%D0%BF%D0%B8%D1%8F_(cropped).jpg)
