Микроорганизм может иметь от одного до многих жгутиков. Грамотрицательная бактерия Helicobacter pylori, например, использует свои жгутики, чтобы продвигаться через желудок и достигать слизистой оболочки , где она может колонизировать эпителий и потенциально вызвать гастрит и язву – фактор риска рака желудка . [5] У некоторых роящихся бактерий жгутик может также функционировать как сенсорная органелла , будучи чувствительным к влажности вне клетки. [6]
В трех доменах бактерий , архей и эукариот жгутик имеет разную структуру, белковый состав и механизм движения, но выполняет одну и ту же функцию обеспечения подвижности . Латинское слово flagellum означает « хлыст », описывая его плавательное движение, напоминающее плеть . Жгутик архей называют археллой , чтобы отметить его отличие от жгутика бактерий. [7] [8]
Эукариотические жгутики и реснички одинаковы по строению, но имеют разную длину и функции. [9] Прокариотические бахромки и пили — это меньшие по размеру и более тонкие придатки с разными функциями.
Типы
Прокариотические (бактериальные и архейные) жгутики совершают вращательные движения, а эукариотические жгутики совершают изгибающиеся движения. Прокариотический жгутик использует вращающийся двигатель , а эукариотический жгутик использует сложную систему скользящих нитей. Эукариотические жгутики управляются АТФ, тогда как прокариотические жгутики могут управляться АТФ (археи) или протонами (бактерии). [10]
Три типа жгутиков: бактериальные, архейные и эукариотические.
Жгутики эукариот имеют динеин и микротрубочки , которые движутся с помощью изгибающего механизма. Бактерии и археи не имеют в жгутиках динеина и микротрубочек и передвигаются с помощью вращательного механизма. [11]
Архейные жгутики ( archaella ) внешне похожи на бактериальные жгутики тем, что также имеют вращающийся двигатель, но отличаются во многих деталях и считаются негомологичными . [17] [18] [19]
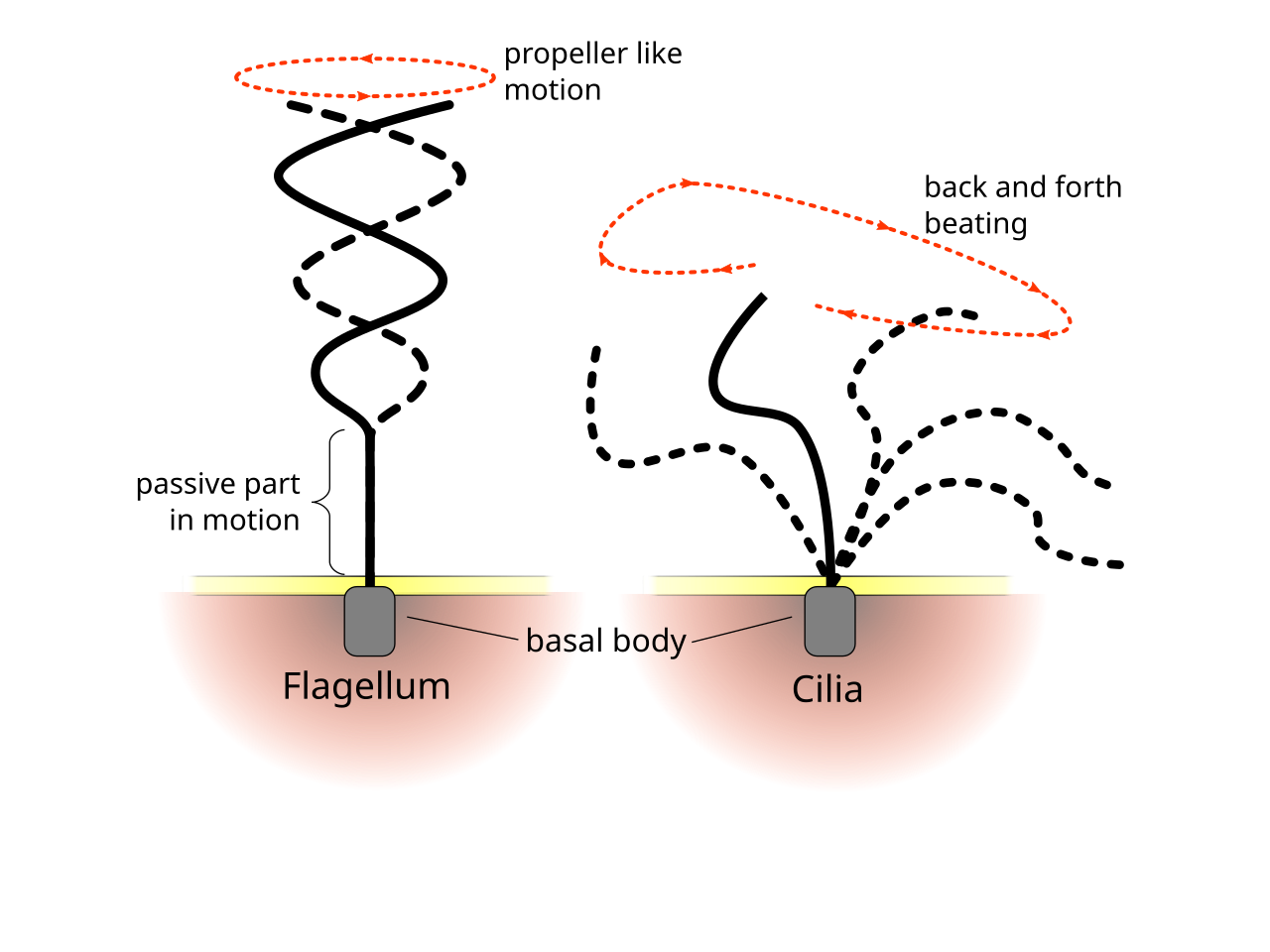
Эукариотические жгутики — жгутики животных, растений и простейших — представляют собой сложные клеточные отростки, которые перемещаются вперед и назад. Эукариотические жгутики и подвижные реснички идентичны по строению, но имеют разную длину, форму волны и функции. Первичные реснички неподвижны и имеют структурно другую аксонему 9+0, а не аксонему 9+2, обнаруженную как в жгутиках, так и в подвижных ресничках.
Жгутиковая нить представляет собой длинный спиральный винт, который продвигает бактерию при вращении двигателя через крючок. У большинства изученных бактерий, включая грамотрицательные Escherichia coli , Salmonella typhimurium , Caulobacter crescentus и Vibrio alginolyticus , нить состоит из 11 протофиламентов, приблизительно параллельных оси нити. Каждый протофиламент представляет собой серию тандемных белковых цепей. Однако Campylobacter jejuni имеет семь протофиламентов. [22]
Базальное тело имеет несколько общих черт с некоторыми типами секреторных пор , например, полая стержнеобразная «пробка» в их центрах, выступающая через плазматическую мембрану. Сходство между бактериальными жгутиками и структурами и белками бактериальной секреторной системы предоставляет научные доказательства, подтверждающие теорию о том, что бактериальные жгутики произошли от системы секреции третьего типа (TTSS).
Атомная структура как бактериальных жгутиков, так и инъектисомы TTSS была выяснена очень подробно, особенно с развитием криоэлектронной микроскопии . Наиболее изученными частями являются части между внутренней и внешней мембраной , то есть каркасные кольца внутренней мембраны (IM), каркасные пары внешней мембраны (OM) и стержень/игла (инъекционома) или стержень/ крючковидные (жгутиковые) отделы. [23]
Мотор
Бактериальный жгутик приводится в движение роторным двигателем ( комплексом Мот ), состоящим из белка, расположенным в точке крепления жгутика на внутренней клеточной мембране. Двигатель приводится в действие за счет протондвижущей силы , т. е. за счет потока протонов (ионов водорода) через мембрану бактериальной клетки за счет градиента концентрации, создаваемого метаболизмом клетки ( виды вибрионов имеют два вида жгутиков: латеральные и полярные, а некоторые из них приводятся в действие ионным насосом натрия , а не протонным насосом [24] ). Ротор переносит протоны через мембрану и при этом вращается. Один только ротор может работать со скоростью от 6000 до 17 000 об/мин , но с прикрепленной жгутиковой нитью обычно скорость достигает только 200–1000 об/мин. Направление вращения может быть изменено переключателем жгутикового двигателя почти мгновенно, что вызвано небольшим изменением положения белка FliG в роторе. [25] Жгутик очень энергоэффективен и потребляет очень мало энергии. [26] [ ненадежный источник? ] Точный механизм генерации крутящего момента до сих пор плохо изучен. [27] Поскольку жгутиковый мотор не имеет выключателя, белок epsE используется в качестве механического сцепления для отключения мотора от ротора, тем самым останавливая жгутик и позволяя бактерии оставаться на одном месте. [28]
Цилиндрическая форма жгутиков подходит для передвижения микроскопических организмов; эти организмы действуют при низком числе Рейнольдса , где вязкость окружающей воды гораздо важнее, чем ее масса или инерция. [29]
Скорость вращения жгутиков меняется в зависимости от интенсивности протондвижущей силы, тем самым обеспечивая определенные формы контроля скорости, а также позволяя некоторым типам бактерий достигать поразительных скоростей, пропорциональных их размеру; некоторые достигают примерно 60 длин клеток в секунду. При такой скорости бактерии потребуется около 245 дней, чтобы преодолеть 1 км; хотя это может показаться медленным, перспектива меняется, когда вводится понятие масштаба. По сравнению с макроскопическими формами жизни, оно действительно очень быстрое, если выражать его в количестве длин тела в секунду. Гепард, например, достигает длины тела всего около 25 раз в секунду. [30]
Используя свои жгутики, бактерии могут быстро перемещаться к аттрактантам и от репеллентов посредством смещенного случайного блуждания , при этом бег и кувыркание вызываются вращением жгутика против и по часовой стрелке соответственно. Два направления вращения не идентичны (относительно движения жгутика) и выбираются молекулярным переключателем. [31]
Сборка
Во время сборки жгутика компоненты жгутика проходят через полые ядра базального тельца и образующуюся нить. Во время сборки белковые компоненты добавляются на кончике жгутика, а не у основания. [32] In vitro жгутиковые нити спонтанно собираются в растворе, содержащем очищенный флагеллин в качестве единственного белка. [33]
Эволюция
По крайней мере, 10 белковых компонентов бактериального жгутика имеют гомологичные белки с системой секреции третьего типа (T3SS), обнаруженной у многих грамотрицательных бактерий [34] , следовательно, один, вероятно, произошел от другого. Поскольку T3SS имеет такое же количество компонентов, что и жгутиковый аппарат (около 25 белков), определить, какой из них развился первым, сложно. Однако жгутиковая система, по-видимому, в целом включает в себя больше белков, включая различные регуляторы и шапероны, поэтому утверждается, что жгутики произошли от T3SS. Однако было также высказано предположение [35] , что жгутик мог развиться первым или обе структуры развивались параллельно. Потребность ранних одноклеточных организмов в подвижности (мобильности) подтверждает, что в ходе эволюции сначала будут выбраны более подвижные жгутики [35] , но T3SS, развивающийся из жгутика, можно рассматривать как «редуктивную эволюцию» и не получает топологической поддержки со стороны филогенетические деревья . [36] Гипотеза о том, что эти две структуры развились отдельно от общего предка, объясняет сходство белков между двумя структурами, а также их функциональное разнообразие. [37]
Жгутики и дебаты о разумном замысле
Некоторые авторы утверждают, что жгутики не могли эволюционировать, полагая, что они могут функционировать должным образом только тогда, когда все белки находятся на месте. Другими словами, жгутиковый аппарат « неуменьшаемо сложен ». [38] Однако многие белки могут быть удалены или мутированы, а жгутик все еще работает, хотя иногда и с пониженной эффективностью. [39] Более того, поскольку многие белки уникальны для определенного числа видов, разнообразие состава жгутиков бактерий было выше, чем ожидалось. [40] Следовательно, жгутиковый аппарат явно очень гибок с эволюционной точки зрения и прекрасно способен терять или приобретать белковые компоненты. Например, обнаружен ряд мутаций, повышающих подвижность кишечной палочки . [41] Дополнительные доказательства эволюции бактериальных жгутиков включают существование рудиментарных жгутиков, промежуточных форм жгутиков и закономерностей сходства между последовательностями жгутиковых белков, включая наблюдение, что почти все основные жгутиковые белки имеют известную гомологию с нежгутиковыми белками. . [34] Кроме того, было установлено, что несколько процессов играют важную роль в эволюции жгутиков, включая самосборку простых повторяющихся субъединиц, дупликацию генов с последующей дивергенцией, рекрутирование элементов из других систем («молекулярный бриколаж») и рекомбинацию. [42]
Жгутиковые механизмы
Различные виды бактерий имеют разное количество и расположение жгутиков, [43] [44] названных термином трихо , от греческого трихос , означающего волосы . [45]
Монотриховые бактерии, такие как Vibrio cholerae, имеют один полярный жгутик . [46]
Амфитриховые бактерии имеют по одному жгутику на каждом из двух противоположных концов (например, Alcaligenes faecalis ) — одновременно действует только один жгутик, что позволяет бактерии быстро изменить курс, переключая какой жгутик активен.
Бактерии Lophotrichous ( греческий термин lopho , означающий гребень или пучок ) [47] имеют множество жгутиков, расположенных в одном и том же месте на бактериальной поверхности, как, например, Helicobacter pylori , которые действуют согласованно, направляя бактерии в одном направлении. Во многих случаях основания множества жгутиков окружены специализированной областью клеточной мембраны, называемой полярной органеллой . [ нужна цитата ]
Перитриховые бактерии имеют жгутики, выступающие во всех направлениях (например, E. coli ).
Вращение монотрихового полярного жгутика против часовой стрелки толкает клетку вперед, при этом жгутик тянется позади, подобно штопору, движущемуся внутри пробки. Вода в микроскопическом масштабе очень вязкая , в отличие от обычной воды .
Спирохеты , напротив, имеют жгутики, называемые эндофлагеллами , отходящие от противоположных полюсов клетки и расположенные внутри периплазматического пространства , как показано путем разрыва внешней мембраны, а также с помощью электронной криотомографической микроскопии. [48] [49] [50] Вращение нитей относительно тела клетки заставляет всю бактерию двигаться вперед по спиралевидному движению, даже через материал, достаточно вязкий, чтобы предотвратить прохождение обычно жгутиковых бактерий.
У некоторых крупных форм Selenomonas более 30 отдельных жгутиков расположены вне тела клетки, спирально переплетаясь друг с другом, образуя толстую структуру (легко видимую в световой микроскоп), называемую «пучком».
У некоторых Vibrio spp. (особенно Vibrio parahaemolyticus [51] ) и родственных бактерий , таких как Aeromonas , две жгутиковые системы сосуществуют, используя разные наборы генов и разные ионные градиенты для получения энергии. Полярные жгутики конститутивно выражены и обеспечивают подвижность в объеме жидкости, тогда как латеральные жгутики выражены, когда полярные жгутики испытывают слишком большое сопротивление при повороте. [52] [53] [54] [55] [56] [57] Они обеспечивают подвижность на поверхностях или в вязких жидкостях.
Объединение
Связывание — это событие, которое может происходить в многожгутиковых клетках, связывая жгутики вместе и заставляя их скоординировано вращаться.
Жгутики представляют собой левосторонние спирали, и при вращении их роторов против часовой стрелки они могут связываться и вращаться вместе. Когда роторы меняют направление, вращаясь по часовой стрелке, жгутик разматывается из пучка. Это может привести к тому, что клетка прекратит свое движение вперед и вместо этого начнет дергаться на месте, что называется кувырком . Переворот приводит к стохастической переориентации клетки, заставляя ее менять направление своего движения вперед.
Неизвестно, какие стимулы приводят к переключению между связыванием и переворачиванием, но двигатель хорошо адаптируется к различным сигналам. В модели, описывающей хемотаксис («намеренное движение»), вращение жгутика по часовой стрелке подавляется химическими соединениями, благоприятными для клетки (например, пищей). При движении в благоприятном направлении концентрация таких химических аттрактантов увеличивается, и поэтому кувырки постоянно подавляются, позволяя двигаться вперед; аналогично, когда направление движения клетки неблагоприятно (например, вдали от химического аттрактанта), кувырки больше не подавляются и происходят гораздо чаще, с вероятностью, что клетка будет таким образом переориентирована в правильном направлении.
Однако даже если бы все жгутики вращались по часовой стрелке, они часто не могли образовать пучок по геометрическим и гидродинамическим причинам. [58] [59]
Эукариотический
Эукариотические жгутики. 1 – аксонема, 2 – клеточная мембрана, 3 – IFT (внутрижгутиковый транспорт), 4 – базальное тельце, 5 – поперечный срез жгутиков, 6 – тройки микротрубочек базального тельцаПоперечное сечение аксонемыПродольный срез области жгутиков Chlamydomonas Reinhardtii . В верхушке клетки находится базальное тельце, служащее местом крепления жгутика. Базальные тельца происходят из центриолей и имеют субструктуру, аналогичную субструктуре центриолей, с девятью триплетами периферических микротрубочек (см. структуру внизу в центре изображения).Структура «9+2» видна на этой микрофотографии поперечного сечения аксонемы.
Терминология
Стремясь подчеркнуть различие между бактериальными жгутиками и эукариотическими ресничками и жгутиками, некоторые авторы пытались заменить название этих двух эукариотических структур на « ундулиподии » (например, все работы Маргулиса с 1970-х годов) [60] или «реснички». для обоих (например, Hülsmann, 1992; [61] Adl et al., 2012; [62] большинство статей Кавальера-Смита ), сохраняя «жгутики» для бактериальной структуры. Однако дискриминационное использование терминов «реснички» и «жгутики» для эукариот, принятое в этой статье (см. § Жгутики в сравнении с ресничками ниже), все еще распространено (например, Andersen et al., 1991; [63] Leadbeater et al., 2000). [64]
Внутренняя структура
Ядро эукариотического жгутика, известное как аксонема , представляет собой пучок из девяти слитых пар микротрубочек , известных как дублеты , окружающих две центральные одиночные микротрубочки ( синглеты ). Эта аксонема 9+2 характерна для жгутика эукариот. В основании эукариотического жгутика находится базальное тельце , «блефаропласт» или кинетосома, которое является центром организации микротрубочек для жгутиковых микротрубочек и имеет длину около 500 нанометров. Базальные тельца структурно идентичны центриолям . Жгутик заключен в плазматическую мембрану клетки , так что внутренняя часть жгутика доступна для цитоплазмы клетки .
Помимо аксонемы и базального тельца, относительно постоянных по морфологии, другими внутренними структурами жгутикового аппарата являются переходная зона (место соединения аксонемы и базального тельца) и корневая система (микротрубчатые или фибриллярные структуры, которые простираются от базальных тел в цитоплазму). ), более изменчивы и полезны в качестве индикаторов филогенетических взаимоотношений эукариот. Другими структурами, более редкими, являются парафгеллярный (или параксиальный, параксонемальный) стержень, R-волокно и S-волокно. [65] : 63–84 О поверхностных структурах см. ниже.
Механизм
Каждая из 9 внешних дублетных микротрубочек простирает пару динеиновых плеч («внутреннее» и «внешнее» плечо) к соседней микротрубочке; они производят силу за счет гидролиза АТФ. Жгутиковая аксонема также содержит радиальные спицы , полипептидные комплексы, простирающиеся от каждого из девяти внешних дублетов микротрубочек к центральной паре, причем «головка» спицы обращена внутрь. Считается, что радиальная спица участвует в регуляции движения жгутиков, хотя ее точная функция и метод действия еще не изучены. [66]
Жгутики против ресничек
Схема биения эукариотических «жгутика» и «циллюма» - традиционное различие до того, как известны структуры этих двух.
Регулярные ритмы ресничек и жгутиков эукариот вызывают движение на клеточном уровне. Примеры варьируются от движения отдельных клеток, например, плавания сперматозоидов , до транспортировки жидкости вдоль стационарного слоя клеток, например, в дыхательных путях . [67]
Хотя эукариотические реснички и жгутики в конечном итоге одинаковы, их иногда классифицируют по характеру движения - традиция, существовавшая еще до того, как были известны их структуры. В случае жгутиков движение чаще плоское и волнообразное, тогда как подвижные реснички часто совершают более сложное трехмерное движение с силовым и восстановительным ходом. [67] Еще одна традиционная форма различия – это количество 9+2 органелл в клетке. [66]
Внутрижгутиковый транспорт
Внутрижгутиковый транспорт , процесс, посредством которого субъединицы аксонемы, трансмембранные рецепторы и другие белки перемещаются вверх и вниз по длине жгутика, важен для правильного функционирования жгутика как в подвижности, так и в передаче сигнала. [68]
Эволюция и возникновение
Эукариотические жгутики или реснички, вероятно, наследственная характеристика, [69] широко распространены почти во всех группах эукариот как относительно постоянное состояние или как стадия жизненного цикла со жгутиками (например, зоиды , гаметы , зооспоры , которые могут производиться постоянно или нет). [70] [71] [62]
Первая ситуация встречается либо в специализированных клетках многоклеточных организмов (например, хоаноцитах губок , либо в мерцательном эпителии многоклеточных животных ), как у инфузорий и многих эукариот с «состоянием жгутиковых» (или «монадоидным уровнем организации», см. Flagellata , искусственная группа).
Для характеристики эукариотов используется ряд терминов, связанных с жгутиками или ресничками. [71] [74] [65] : 60–63 [75] [76] В зависимости от структуры поверхности жгутики могут быть:
хлыстовые жгутики (= гладкие акронематические жгутики): без волосков, например, у Opisthokonta
волосатые жгутики (= мишура, флиммер, плевронематические жгутики): с волосками (= mastigonemes sensu lato ), разделенные на:
трехсторонние (= страминипилусные) волосы: с тремя участками (основание, трубчатый стержень и один или несколько концевых волосков). Встречается в большинстве Гетероконта .
стихонематические жгутики: с одним рядом волосков.
пантонематические жгутики: с двумя рядами волосков.
акронематические: жгутики с одиночной терминальной мастигонемой или жгутиковыми волосками (например, бодониды ); [77] некоторые авторы используют этот термин как синоним хлыстовой травмы.
опистоконт: клетки со жгутиками, прикрепленными сзади, например у Opisthokonta (Vischer, 1945). У Haptophyceae жгутики прикреплены латерально или терминально, но во время быстрого плавания направлены назад. [81]
акроконт: клетки со жгутиками, прикрепленными апикально.
субакроконт: клетки со жгутиками, вставленными субапикально.
плевроконт: клетки со жгутиками, вставленными латерально.
По схеме избиения:
скольжение: жгутик, тянущийся по субстрату [78]
гетеродинамические: жгутики с разным характером биения (обычно один жгутик функционирует при захвате пищи, а другой - при скольжении, закреплении, движении или «управлении») [82]
изодинамические: биение жгутиков по одному и тому же принципу.
Другие термины, относящиеся к жгутиковому типу:
изоконт: клетки со жгутиками одинаковой длины. Раньше оно также использовалось для обозначения Chlorophyta .
гетероконт: термин, введенный Лютером (1899) для обозначения Xanthophyceae из -за пары жгутиков неравной длины. Особый смысл оно приобрело применительно к клеткам с передним страминипилозным жгутиком (с трехраздельными мастигонемами, расположенными в один или два ряда) и задним, обычно гладким жгутиком. Он также используется для обозначения таксона Heterokonta .
стефаноконт: клетки с венцом жгутиков вблизи переднего конца, например, гаметы и споры Oedogoniales , споры некоторых Bryopsidales . Термин, введенный Блэкманом и Тэнсли (1902) для обозначения Oedogoniales .
аконт: клетки без жгутиков. Он также использовался для обозначения таксономических групп, таких как Aconta или Akonta: Zygnematophyceae и Bacillariophyceae (Oltmanns, 1904) или Rhodophyceae (Christensen, 1962).
Архейный
Архееллум некоторых видов архей внешне похож на жгутик бактерий; в 1980-х годах их считали гомологичными на основании общей морфологии и поведения. [83] И жгутики, и архаеллы состоят из нитей, выходящих за пределы клетки и вращающихся, приводя в движение клетку. Жгутики архей имеют уникальную структуру, в которой отсутствует центральный канал. Подобно бактериальным пилинам IV типа , архейные белки (архееллины) состоят из сигнальных пептидов класса 3 и процессируются ферментом, подобным препилиновой пептидазе IV типа. Архееллины обычно модифицируются путем добавления N-связанных гликанов , которые необходимы для правильной сборки или функционирования. [3]
Открытия 1990-х годов выявили многочисленные детальные различия между жгутиками архей и бактерий. К ним относятся:
Вращение жгутиков бактерий осуществляется за счет движущей силы протонов – потока ионов Н + или изредка движущей силы натрия – потока ионов Na + ; Вращение жгутиков архей осуществляется за счет АТФ . [84]
В то время как бактериальные клетки часто имеют множество жгутиковых нитей, каждая из которых вращается независимо, жгутик архей состоит из пучка множества нитей, которые вращаются как единое целое.
Бактериальные жгутики растут за счет добавления субъединиц флагеллина на кончике; жгутики архей растут за счет присоединения субъединиц к основанию.
Бактериальные жгутики толще, чем архаеллы, и бактериальная нить имеет внутри достаточно большую полую «трубку», благодаря которой субъединицы флагеллина могут течь вверх по внутренней части нити и добавляться на кончике; архаеллум слишком тонкий (12-15 нм), чтобы это позволить. [85]
Многие компоненты бактериальных жгутиков имеют сходство последовательностей с компонентами систем секреции типа III , но компоненты бактериальных жгутиков и архаелл не имеют сходства последовательностей. Вместо этого некоторые компоненты архаелл имеют общую последовательность и морфологическое сходство с компонентами пилей типа IV , которые собираются под действием систем секреции типа II (номенклатура пилей и систем секреции белка не согласована). [85]
Эти различия подтверждают теорию о том, что бактериальные жгутики и архаеллы являются классическим случаем биологической аналогии или конвергентной эволюции , а не гомологии . [86] [87] [88] Исследования структуры архаелл достигли значительного прогресса, начиная с начала 2010-х годов, с появлением первой структуры с атомным разрешением белка архаеллы, открытием дополнительных функций архаеллы и первыми сообщениями об архаеллах в Наноархеота и Таумархеота. [89] [90]
Грибковый
Единственные грибы , у которых на спорах имеется один жгутик, — это хитриды . У Batrachochytrium dendrobatidis длина жгутика составляет 19–20 мкм. [91] Нефункционирующая центриоль лежит рядом с кинетосомой . Девять связанных между собой опор прикрепляют кинетосому к плазмалемме , а в переходной зоне имеется терминальная пластинка. На поперечном срезе наблюдалась внутренняя кольцеобразная структура, прикрепленная к канальцам дублетов жгутиков внутри переходной зоны. [91]
Дополнительные изображения
Множественные жгутики в лофотриховом расположении на поверхности Helicobacter pylori
^ Silflow CD, Лефевр, Пенсильвания (декабрь 2001 г.). «Сборка и подвижность эукариотических ресничек и жгутиков. Уроки Chlamydomonas Reinhardtii». Физиология растений . 127 (4): 1500–7. дои : 10.1104/стр.010807. ПМК 1540183 . ПМИД 11743094.
^ Аб Джаррелл К., изд. (2009). Пили и жгутики: текущие исследования и будущие тенденции . Кайстер Академик Пресс. ISBN978-1-904455-48-6.
^ Мало А.Ф., Гомендио М., Гард Дж., Ланг-Лентон Б., Солер А.Дж., Ролдан Э.Р. (июнь 2006 г.). «Дизайн спермы и функция спермы». Письма по биологии . 2 (2): 246–9. дои : 10.1098/rsbl.2006.0449. ПМЦ 1618917 . ПМИД 17148374.
^ Лейси Б.Е., Роузмор Дж. (октябрь 2001 г.). «Helicobacter pylori: язвы и многое другое: начало эпохи». Журнал питания . 131 (10): 2789С–2793С. дои : 10.1093/jn/131.10.2789S . PMID 11584108. Архивировано из оригинала (аннотация страницы) 7 февраля 2009 года . Проверено 2 июня 2008 г.
^ Ван Кью, Сузуки А, Мариконда С, Порволлик С, Харши Р.М. (июнь 2005 г.). «Ощущение влажности: новая роль бактериального жгутика». Журнал ЭМБО . 24 (11): 2034–42. дои : 10.1038/sj.emboj.7600668. ПМЦ 1142604 . ПМИД 15889148.
↑ Альберс С.В., Джаррелл К.Ф. (27 января 2015 г.). «Архееллум: как плавают археи». Границы микробиологии . 6:23 . дои : 10.3389/fmicb.2015.00023 . ПМЦ 4307647 . ПМИД 25699024.
^ Квакс, ТЭФ; Альберс, СВ; Пфайффер, Ф (14 декабря 2018 г.). «Такси в архее». Новые темы в науках о жизни . 2 (4): 535–546. doi : 10.1042/ETLS20180089. ПМК 7289035 . ПМИД 33525831.
^ Хаймо LT, Розенбаум JL (декабрь 1981 г.). «Реснички, жгутики и микротрубочки». Журнал клеточной биологии . 91 (3, часть 2): 125–130 с. дои : 10.1083/jcb.91.3.125s. ПМК 2112827 . ПМИД 6459327.
^ Штрайф С., Штаудингер В.Ф., Марван В., Остерхельт Д. (2008). «Вращение жгутиков у архей Halobacterium salinarum зависит от АТФ». Журнал молекулярной биологии . 384 (1): 1–8. дои : 10.1016/j.jmb.2008.08.057. ПМИД 18786541.
^ аб Альбертс, Брюс (2015). Молекулярная биология клетки (Шестое изд.). Нью-Йорк, штат Нью-Йорк. п. 942. ИСБН9780815344643.{{cite book}}: CS1 maint: location missing publisher (link)
^ Сильверман М., Саймон М. (май 1974 г.). «Вращение жгутиков и механизм подвижности бактерий». Природа . 249 (452): 73–4. Бибкод : 1974Natur.249...73S. дои : 10.1038/249073a0. PMID 4598030. S2CID 10370084.
^ Ян Т.Л., Бове EC (1965). «Передвижение и локомоция микроорганизмов». Ежегодный обзор микробиологии . 19 :21–58. doi : 10.1146/annurev.mi.19.100165.000321. ПМИД 5318439.
^ Харши РМ (2003). «Подвижность бактерий на поверхности: много путей к общей цели». Ежегодный обзор микробиологии . 57 : 249–73. doi : 10.1146/annurev.micro.57.030502.091014. ПМИД 14527279.
^ Нг С.Ю., Чабан Б., Джаррелл К.Ф. (2006). «Архейные жгутики, бактериальные жгутики и пили IV типа: сравнение генов и посттрансляционных модификаций». Журнал молекулярной микробиологии и биотехнологии . 11 (3–5): 167–91. дои : 10.1159/000094053. PMID 16983194. S2CID 30386932.
^ Метлина А.Л. (ноябрь 2004 г.). «Бактериальные и архейные жгутики как органеллы подвижности прокариот». Биохимия. Биохимия . 69 (11): 1203–12. дои : 10.1007/s10541-005-0065-8. PMID 15627373. S2CID 632440.
^ Джаррелл К. (2009). «Археальные жгутики и пили». Пили и жгутики: текущие исследования и будущие тенденции . Кайстер Академик Пресс. ISBN978-1-904455-48-6.
^ Уорролл, Лиам Дж.; Маевски, Дороти Д.; Стринадка, Наталья CJ (15 сентября 2023 г.). «Структурные данные о системах секреции типа III бактериального жгутика и инъекциономы». Ежегодный обзор микробиологии . 77 (1): 669–698. doi : 10.1146/annurev-micro-032521-025503 . ISSN 0066-4227. PMID 37713458. S2CID 261963968.
^ Ацуми Т., Маккартер Л., Имаэ Ю. (январь 1992 г.). «Полярные и латеральные жгутиковые моторы морских вибрионов приводятся в движение разными ионно-движущими силами». Природа . 355 (6356): 182–4. Бибкод : 1992Natur.355..182A. дои : 10.1038/355182a0. PMID 1309599. S2CID 4315167.
^ Дин Т (2 августа 2010 г.). «Внутри самого эффективного двигателя природы: жгутика». Австралийский учёный-биолог .
^ Нагата Ю (июнь 2014 г.). «Раскрытие секретов природного наномотора». Азиатский обзор Nikkei .
^ Мора Т., Ю Х., Сова Ю., Вингрин Н.С. (октябрь 2009 г.). «Этапы бактериального жгутикового мотора». PLOS Вычислительная биология . 5 (10): е1000540. arXiv : 0904.0438 . Бибкод : 2009PLSCB...5E0540M. дои : 10.1371/journal.pcbi.1000540 . ПМК 2759076 . ПМИД 19851449.
^ Уитфилд Дж. (19 июня 2008 г.). «Бактериальные двигатели имеют свое сцепление». Новости природы : новости.2008.903. дои : 10.1038/news.2008.903 . Проверено 17 мая 2017 г.
^ Дюсенбери БД (2009). «Глава 13». Жизнь в микромасштабе: неожиданная физика маленького размера . Кембридж: Издательство Гарвардского университета. ISBN978-0-674-03116-6.
^ Хильдебранд М (ноябрь 1959 г.). «Движения бегущего гепарда и лошади». Журнал маммологии . 44 (4): 481–495. дои : 10.2307/1376265. JSTOR 1376265.Хотя, по словам Хантера, Люка; Хамман, Дэйв (2003). Гепард . Издательство Струик. стр. 37–38. Самая быстрая зарегистрированная скорость гепарда составила 110 км/ч (68 миль в час).
^ Медоуз Р. (май 2011 г.). «Как бактерии переключают передачи». ПЛОС Биология . 9 (5): e1001061. дои : 10.1371/journal.pbio.1001061 . ПМК 3091840 . ПМИД 21572986.
^ Минамино Т., Имада К., Намба К. (ноябрь 2008 г.). «Механизмы экспорта белков типа III для сборки жгутиков бактерий». Молекулярные биосистемы . 4 (11): 1105–15. дои : 10.1039/b808065h. ПМИД 18931786.
^ Асакура С., Эгучи Г., Иино Т. (октябрь 1964 г.). «Восстановление бактериальных жгутиков in vitro». Журнал молекулярной биологии . 10 : 42–56. дои : 10.1016/S0022-2836(64)80026-7. ПМИД 14222895.
^ аб Паллен MJ, Мацке, штат Нью-Джерси (октябрь 2006 г.). «От происхождения видов к происхождению бактериальных жгутиков». Обзоры природы. Микробиология . 4 (10): 784–90. doi : 10.1038/nrmicro1493. PMID 16953248. S2CID 24057949.
^ ab Saier MH (март 2004 г.). «Эволюция систем секреции белков бактериального типа III». Тенденции в микробиологии . 12 (3): 113–5. дои : 10.1016/j.tim.2004.01.003. ПМИД 15001186.
^ Гофна У, Рон Э.З., Граур Д. (июль 2003 г.). «Бактериальные системы секреции типа III являются древними и развиваются в результате множественных событий горизонтального переноса». Джин . 312 : 151–63. дои : 10.1016/S0378-1119(03)00612-7. ПМИД 12909351.
^ Макканн ХК, Гуттман Д.С. (2008). «Эволюция системы секреции типа III и ее эффекторов во взаимодействиях растений и микробов». Новый фитолог . 177 (1): 33–47. дои : 10.1111/J.1469-8137.2007.02293.X . ПМИД 18078471.
^ Бехе, М. (2007) Грань эволюции. Свободная пресса, Нью-Йорк
^ Раджагопала С.В., Титц Б., Голл Дж., Пэрриш Дж.Р., Уолболд К., Маккевитт М.Т., Палцкилл Т., Мори Х., Финли Р.Л., Уетц П. (2007). «Белковая сеть подвижности бактерий». Молекулярная системная биология . 3 : 128. дои : 10.1038/msb4100166. ЧВК 1943423 . ПМИД 17667950.
^ Титц Б., Раджагопала С.В., Эстер С., Хойзер Р., Утц П. (ноябрь 2006 г.). «Новый консервативный фактор сборки бактериального жгутика». Журнал бактериологии . 188 (21): 7700–6. дои : 10.1128/JB.00820-06. ПМЦ 1636259 . ПМИД 16936039.
^ Какканат А., Фан, доктор медицинских наук, Ло А.В., Битсон С.А., Шембри Массачусетс (10 мая 2017 г.). «Новые гены, связанные с повышенной подвижностью Escherichia coli ST131». ПЛОС ОДИН . 12 (5): e0176290. Бибкод : 2017PLoSO..1276290K. дои : 10.1371/journal.pone.0176290 . ПМК 5425062 . ПМИД 28489862.
^ Паллен М.Дж., Гофна У (2005). «Бактериальные жгутики и секреция типа III: тематические исследования эволюции сложности». Геномная динамика . 3 : 30–47. дои : 10.1159/000107602. ISBN978-3-8055-8340-4. ПМИД 18753783.
^ «Бактериальные жгутики» (PDF) . Архивировано (PDF) из оригинала 9 октября 2022 года . Проверено 29 декабря 2021 г.
^ Руан Дж, Като Т, Сантини CL, Мията Т, Кавамото А, Чжан В.Дж., Бернадак А, Ву Л.Ф., Намба К. (декабрь 2012 г.). «Архитектура жгутикового аппарата быстроплавающей магнитотаксической бактерии МО-1». Proc Natl Acad Sci США . 109 (50): 20643–8. Бибкод : 2012PNAS..10920643R. дои : 10.1073/pnas.1215274109 . ПМЦ 3528567 . ПМИД 23184985.
^ "трихо-префикс" . Проверено 26 марта 2022 г.
^ Эчазаррета, Массачусетс; Клозе, Кентукки (2019). «Синтез жгутиков вибрионов». Границы клеточной и инфекционной микробиологии . 9 : 131. дои : 10.3389/fcimb.2019.00131 . ПМК 6504787 . ПМИД 31119103.
^ "Лофо" . Проверено 26 марта 2022 г.
^ Изард Дж., Ренкен С., Се CE, Дерозье, округ Колумбия, Данэм-Эмс С., Ла Ваке С., Гебхардт Л.Л., Лимбергер Р.Дж., Кокс Д.Л., Марко М., Радольф Дж.Д. (декабрь 2009 г.). «Криоэлектронная томография поясняет молекулярную архитектуру Treponema pallidum, спирохеты сифилиса». Журнал бактериологии . 191 (24): 7566–80. дои : 10.1128/JB.01031-09. ПМЦ 2786590 . ПМИД 19820083.
^ Изард Дж., Се CE, Лимбергер Р.Дж., Маннелла Калифорния, Марко М. (июль 2008 г.). «Нативная клеточная архитектура Treponema denticola, выявленная с помощью криоэлектронной томографии». Журнал структурной биологии . 163 (1): 10–7. дои : 10.1016/j.jsb.2008.03.009. ПМЦ 2519799 . ПМИД 18468917.
^ Кудряшев М., Цирклафф М., Баумайстер В., Саймон М.М., Валлих Р., Фришкнехт Ф. (март 2009 г.). «Сравнительная криоэлектронная томография патогенных спирохет болезни Лайма». Молекулярная микробиология . 71 (6): 1415–34. дои : 10.1111/j.1365-2958.2009.06613.x . PMID 19210619. S2CID 19650892.
^ Ким Ю.К., Маккартер Л.Л. (июль 2000 г.). «Анализ полярной генной системы жгутиков Vibrio parahaemolyticus». Журнал бактериологии . 182 (13): 3693–704. дои : 10.1128/JB.182.13.3693-3704.2000. ПМК 94540 . ПМИД 10850984.
^ Ацуми Т., Маэкава Ю., Ямада Т., Кавагиши И., Имаэ Ю., Хомма М. (август 1996 г.). «Влияние вязкости на плавание боковых и полярных жгутиков Vibrio alginolyticus». Журнал бактериологии . 178 (16): 5024–6. дои : 10.1128/jb.178.16.5024-5026.1996. ПМК 178290 . ПМИД 8759871.
^ Маккартер Л.Л. (2004). «Двойная жгутиковая система обеспечивает подвижность при разных обстоятельствах». Журнал молекулярной микробиологии и биотехнологии . 7 (1–2): 18–29. дои : 10.1159/000077866. PMID 15170400. S2CID 21963003.
^ Мерино С., Шоу Дж.Г., Томас Дж.М. (октябрь 2006 г.). «Бактериальные боковые жгутики: индуцируемая система жгутиков». Письма FEMS по микробиологии . 263 (2): 127–35. дои : 10.1111/j.1574-6968.2006.00403.x . ПМИД 16978346.
^ Белас Р., Саймон М., Сильверман М. (июль 1986 г.). «Регуляция транскрипции гена боковых жгутиков у Vibrio parahaemolyticus». Журнал бактериологии . 167 (1): 210–8. дои : 10.1128/jb.167.1.210-218.1986. ПМК 212863 . ПМИД 3013835.
^ Каналс Р., Алтарриба М., Вилчес С., Хорсбург Г., Шоу Дж.Г., Томас Дж.М., Мерино С. (февраль 2006 г.). «Анализ генной системы латеральных жгутиков Aeromonas Hydrophila AH-3». Журнал бактериологии . 188 (3): 852–62. дои : 10.1128/JB.188.3.852-862.2006. ПМЦ 1347325 . ПМИД 16428388.
^ Каналс Р., Рамирес С., Вилчес С., Хорсбург Г., Шоу Дж.Г., Томас Дж.М., Мерино С. (январь 2006 г.). «Биогенез полярных жгутиков у Aeromonas Hydrophila». Журнал бактериологии . 188 (2): 542–55. дои : 10.1128/JB.188.2.542-555.2006. ПМЦ 1347287 . ПМИД 16385045.
^ Ким М., Бёрд Дж.К., Ван Парис А.Дж., Брейер К.С., Пауэрс Т.Р. (декабрь 2003 г.). «Макроскопическая модель объединения жгутиков бактерий». Труды Национальной академии наук Соединенных Штатов Америки . 100 (26): 15481–5. arXiv : cond-mat/0312562 . Бибкод : 2003PNAS..10015481K. дои : 10.1073/pnas.2633596100 . ПМЦ 307593 . ПМИД 14671319.
^ Macnab RM (январь 1977 г.). «Бактериальные жгутики, вращающиеся в пучках: исследование спиральной геометрии». Труды Национальной академии наук Соединенных Штатов Америки . 74 (1): 221–5. Бибкод : 1977PNAS...74..221M. дои : 10.1073/pnas.74.1.221 . ПМЦ 393230 . ПМИД 264676.
^ Тейлор FJ (ноябрь 2003 г.). «Крах системы двух королевств, подъем протистологии и основание Международного общества эволюционной протистологии (ISEP)». Международный журнал систематической и эволюционной микробиологии . 53 (Часть 6): 1707–14. дои : 10.1099/ijs.0.02587-0 . ПМИД 14657097.
^ Хюльсманн Н. (август 1992 г.). «Ундулиподий: Конец бесполезной дискуссии». Европейский журнал протистологии . 28 (3): 253–7. дои : 10.1016/s0932-4739(11)80231-2. ПМИД 23195228.
^ abc Адл С.М., Симпсон А.Г., Лейн CE, Лукеш Дж., Басс Д., Баузер С.С. и др. (сентябрь 2012 г.). «Пересмотренная классификация эукариот». Журнал эукариотической микробиологии . 59 (5): 429–93. дои : 10.1111/j.1550-7408.2012.00644.x. ПМЦ 3483872 . ПМИД 23020233.
^ Андерсен Р.А., Барр DJ, Линн Д.Х., Мелконян М., Моэструп О., Сани М.А. (1991). «Терминология и номенклатура элементов цитоскелета, связанных с жгутиковым/ресничным аппаратом у простейших». Протоплазма . 164 (1–3): 1–8. дои : 10.1007/bf01320809. S2CID 40755371.
^ Ледбитер, Барри СК; Грин, Джон К., ред. (2000). Жгутиконосцы: единство, разнообразие и эволюция. Специальный том Ассоциации систематики. Том. 59. Тейлор и Фрэнсис. ISBN978-1-4822-6822-5.
^ аб Барсанти Л., Гуальтьери П. (2006). Водоросли: анатомия, биохимия и биотехнология. Флорида, США: CRC Press. ISBN9780203492598.
^ Аб Линдеманн, CB; Лесич, К.А. (15 февраля 2010 г.). «Жгутиковое и цилиарное избиение: доказанное и возможное». Журнал клеточной науки . 123 (Часть 4): 519–28. дои : 10.1242/jcs.051326. PMID 20145000. S2CID 18673550.
^ аб Лодиш, Харви; Берк, Арнольд; Зипурски, С. Лоуренс; Мацудайра, Пол; Балтимор, Дэвид; Дарнелл, Джеймс (2000). «Раздел 19.4 Реснички и жгутики: строение и движение». Реснички и жгутики: строение и движение. ISBN0-7167-3136-3.
^ Пасур GJ (октябрь 2004 г.). «Внутрижгутиковый транспорт и реснично-зависимое заболевание почек: цилиарная гипотеза поликистозной болезни почек». Журнал Американского общества нефрологов . 15 (10): 2528–36. дои : 10.1097/01.ASN.0000141055.57643.E0 . ПМИД 15466257.
^ Юбуки Н., Леандер Б.С. (июль 2013 г.). «Эволюция центров организации микротрубочек на дереве эукариот». Заводской журнал . 75 (2): 230–44. дои : 10.1111/tpj.12145 . ПМИД 23398214.
^ Рэйвен, JA (2000). «Состояние жгутика». Ледбитер и Грин, 2000, стр. 27–48 . ЦРК Пресс. ISBN9781482268225.
^ аб Вебстер Дж., Вебер Р. (25 января 2007 г.). «Споры грибов». 2007 (3-е изд.). Кембридж: Издательство Кембриджского университета. стр. 23–24. ISBN9781139461504.
^ ab Jeuck A, Арндт H (ноябрь 2013 г.). «Краткий справочник по распространенным гетеротрофным жгутиконосцам пресноводных местообитаний, основанный на морфологии живых организмов». Протист . 164 (6): 842–60. дои : 10.1016/j.protis.2013.08.003 . ПМИД 24239731.
^ Сани М (1989). Простейшие и другие протисты. Лондон: Эдвард Арнольд. стр. 98–99. ISBN9780521428057.
^ Воробей ФК (1960). Водные фикомицеты (2-е изд.). Анн-Арбор: Мичиган: Издательство Мичиганского университета. п. 15.
^ Хибберд DJ (1976). «Ультраструктура и таксономия Chrysophyceae и Prymnesiophyceae (Haptophyceae): обзор с некоторыми новыми наблюдениями по ультраструктуре Chrysophyceae». Журнал Лондонского Линнеевского общества, Ботаника . 72 (2): 55–80. doi :10.1111/j.1095-8339.1976.tb01352.x.
^ Кавалер-Смит Т (1987). «Происхождение эукариотических и архебактериальных клеток». Анналы Нью-Йоркской академии наук . 503 (1): 17–54. Бибкод : 1987NYASA.503...17C. doi :10.1111/j.1749-6632.1987.tb40596.x. PMID 3113314. S2CID 38405158.[ постоянная мертвая ссылка ]
^ Мэдиган, Майкл Т. (2019). Брока биология микроорганизмов (Пятнадцатое, Глобальное изд.). Нью-Йорк, Нью-Йорк. стр. 70–71. ISBN9781292235103.{{cite book}}: CS1 maint: location missing publisher (link)
^ Аб Гош А., Альберс С.В. (январь 2011 г.). «Сборка и функция жгутика архей». Труды Биохимического общества . 39 (1): 64–9. дои : 10.1042/BST0390064. PMID 21265748. S2CID 23810797.
^ Томас Н.А., Барди С.Л., Джаррелл К.Ф. (апрель 2001 г.). «Жгутик архей: другой вид структуры подвижности прокариот». Обзоры микробиологии FEMS . 25 (2): 147–74. doi :10.1111/j.1574-6976.2001.tb00575.x. PMID 11250034. S2CID 34411164.
^ Чимилески, Скотт; Папке, Р. Тейн (2015). «Как получить доступ к архейным пилям IV типа: расширяющийся репертуар клеточных придатков предполагает сложную регуляцию и разнообразные функции». Границы микробиологии . 6 : 362. дои : 10.3389/fmicb.2015.00362 . ISSN 1664-302X. ПМЦ 4419858 . ПМИД 25999922.
^ де Соуза Мачадо, Дж. Нуно; Воллмар, Леони; Шимпф, Джулия; Чаудхури, Паушали; Кумария, Рашми; ван дер Дос, Крис; Хьюгель, Торстен; Альберс, Соня-Верена (2021). «Аутофосфорилирование KaiC-подобного белка ArlH ингибирует олигомеризацию и взаимодействие с ArlI, моторной АТФазой археллы». Молекулярная микробиология . 116 (3): 943–956. дои : 10.1111/mmi.14781 . ISSN 0950-382X. ПМИД 34219289.
^ Джаррелл, Кен Ф; Альберс, Соня-Верена; Мачадо, Дж. Нуно де Соуза (2021). «Всесторонняя история подвижности и археляции в археях». ФЭМС Микробы . 2 : xtab002. doi : 10.1093/femsmc/xtab002. ISSN 2633-6685. ПМЦ 10117864 . ПМИД 37334237.
^ ab Longcore JE, Pessier AP, Nichols DK (1999). « Batrachochytrium Dendrobatidis gen. et sp. nov , хитридий, патогенный для амфибий». Микология . 91 (2): 219–227. дои : 10.2307/3761366. JSTOR 3761366.
дальнейшее чтение
Берг ХК (январь 2000 г.). «Подвижное поведение бактерий». Физика сегодня . 53 (1): 24–29. Бибкод : 2000PhT....53a..24B. дои : 10.1063/1.882934 . S2CID 178516210.
Линдеманн С. (4 апреля 2008 г.). «Механизмы подвижности сперматозоидов». Оклендский университет. Архивировано из оригинала 16 мая 2008 года . Проверено 18 мая 2008 г.
Перселл Э.М. (1977). «Жизнь при низком числе Рейнольдса» (PDF) . Американский журнал физики . 45 (1): 3–11. Бибкод : 1977AmJPh..45....3P. дои : 10.1119/1.10903. hdl : 2433/226838 . Архивировано из оригинала (PDF) 5 июня 2011 года . Проверено 19 октября 2009 г.
Мацке, штат Нью-Джерси (10 ноября 2003 г.). «Эволюция в (броуновском) пространстве: модель происхождения бактериального жгутика». talkdesign.org.