Змеи — удлиненные, лишенные конечностей , хищные рептилии подотряда Serpentes ( / s ɜːr ˈ p ɛ n t iː z / ). [2] Как и все другие чешуйчатые , змеи являются экзотермическими позвоночными амниотами , покрытыми перекрывающимися чешуйками . У многих видов змей черепа имеют на несколько больше суставов, чем у их предков -ящериц , что позволяет им заглатывать добычу, размер которой намного превышает размеры их головы ( краниальный кинез ). Чтобы разместить свое узкое тело, парные органы змей (например, почки) располагаются один перед другим, а не рядом, и у большинства из них есть только одно функциональное легкое . У некоторых видов сохраняется тазовый пояс с парой рудиментарных когтей по обе стороны от клоаки . Ящерицы независимо развивали удлиненные тела без конечностей или со значительно уменьшенными конечностями, по крайней мере, двадцать пять раз в результате конвергентной эволюции , что привело к появлению множества линий безногих ящериц . [3] Они напоминают змей, но у некоторых распространенных групп безногих ящериц есть веки и наружные уши, которых нет у змей, хотя это правило не является универсальным (см. Amphisbaenia , Dibamidae и Pygopodidae ).
Живые змеи встречаются на всех континентах, кроме Антарктиды, и на большинстве небольших участков суши; исключения включают некоторые крупные острова, такие как Ирландия, Исландия, Гренландия, Гавайский архипелаг и острова Новой Зеландии, а также множество мелких островов Атлантического и центральной части Тихого океана. [4] Кроме того, морские змеи широко распространены в Индийском и Тихом океанах. В настоящее время известно около тридцати семейств , включающих около 520 родов и около 3900 видов . [5] Их размер варьируется от крошечной барбадосской змеи длиной 10,4 см (4,1 дюйма) [6] до сетчатого питона длиной 6,95 метра (22,8 фута). [7] Ископаемый вид Titanoboa cerrejonensis имел длину 12,8 метра (42 фута). [8] Считается, что змеи произошли от роющих или водных ящериц, возможно, в юрский период, причем самые ранние известные окаменелости датируются периодом от 143 до 167 млн лет назад. [9] [10] Разнообразие современных змей появилось в эпоху палеоцена ( ок. 66–56 млн лет назад, после мел-палеогенового вымирания ). Самые старые сохранившиеся описания змей можно найти в Бруклинском папирусе .
Большинство видов змей неядовиты, а те, у кого есть яд, используют его в первую очередь для убийства и подчинения добычи, а не для самообороны. Некоторые из них обладают ядом, достаточно сильным, чтобы причинить людям болезненные травмы или смерть. Неядовитые змеи либо заглатывают добычу живьем, либо убивают ее сжиманием .
Английское слово « змея» происходит от древнеанглийского snaca , которое само по себе происходит от протогерманского * snak-an- ( ср. германское Schnake «кольцевая змея», шведское snok «травяная змея»), от протоиндоевропейского корня * (s)nēg. -o- 'ползти, чтобы ползти', что также дает подкрадываться , а также санскритское nagá 'змея'. [11] Слово вытеснило гадюку , поскольку гадюка сузила свое значение, хотя в древнеанглийском языке næddre было общим словом, обозначающим змею. [12] Другой термин, змея , происходит от французского языка, в конечном итоге от индоевропейского * serp- «ползти», [13] который также дал древнегреческое ἕρπω ( hérpō ) «Я ползаю» и санскритское sarpá «змея». [ нужна цитата ]
Летопись окаменелостей змей относительно скудна, поскольку скелеты змей обычно маленькие и хрупкие, поэтому окаменелость встречается редко. Окаменелости, легко идентифицируемые как змеи (хотя часто сохраняющие задние конечности), впервые появляются в летописи окаменелостей в меловой период. [15] Самые ранние известные окаменелости настоящих змей (члены коронной группы Serpentes) происходят от морских симолиофиид , самым старым из которых является позднемеловой ( сеноманский возраст) Haasiophis terrasanctus с Западного берега , [1] датируемый между 112 и 112 годами. 94 миллиона лет. [16]
На основании сравнительной анатомии существует мнение, что змеи произошли от ящериц . [17] : 11 [18] Питоны и удавы — примитивные группы современных змей — имеют рудиментарные задние конечности: крошечные когтистые пальцы, известные как анальные шпоры , которые используются для захвата во время спаривания. [17] : 11 [19] Семейства Leptotyphlopidae и Typhlopidae также обладают остатками тазового пояса, которые при видимом виде выглядят как роговые выступы.
Передние конечности отсутствуют у всех известных змей. Это вызвано эволюцией их Hox-генов , контролирующих морфогенез конечностей . Осевой скелет общего предка змей, как и большинство других четвероногих, имел региональную специализацию, состоящую из шейных (шеи), грудных (грудных), поясничных (нижней части спины), крестцовых (тазовых) и хвостовых (хвостовых) позвонков. На ранних этапах эволюции змей экспрессия гена Hox в осевом скелете, ответственная за развитие грудной клетки, стала доминантной. В результате все позвонки перед зачатками задних конечностей (если они есть) имеют одинаковую грудную идентичность (за исключением атланта , оси и 1–3 шейных позвонков). Другими словами, большая часть скелета змеи представляет собой чрезвычайно вытянутую грудную клетку. Ребра находятся исключительно на грудных позвонках. Число шейных, поясничных и тазовых позвонков сильно уменьшено (имеются только 2–10 поясничных и тазовых позвонков), а от хвостовых позвонков остался лишь короткий хвост. Тем не менее, хвост все еще достаточно длинный, чтобы иметь важное применение у многих видов, а у некоторых водных и древесных видов он видоизменился.
Многие современные группы змей возникли в палеоцене , наряду с адаптивной радиацией млекопитающих после вымирания (нептичьих) динозавров . Расширение лугов в Северной Америке также привело к взрывной радиации среди змей. [20] Раньше змеи были второстепенным компонентом североамериканской фауны, но в миоцене число видов и их распространенность резко возросли с первыми появлениями гадюк и элапид в Северной Америке и значительной диверсификацией Colubridae (включая происхождение многих современных родов, таких как Nerodia , Lampropeltis , Pituophis и Pantherophis ). [20]
Существуют ископаемые свидетельства, позволяющие предположить, что змеи могли произойти от роющих ящериц [21] в меловой период . [22] Ранний ископаемый родственник змеи, Наджаш рионегрина , был двуногим роющим животным с крестцом и был полностью наземным . [23] Одним из сохранившихся аналогов этих предполагаемых предков является безухий варан Лантанот с Борнео (хотя он также ведет полуводный образ жизни ). [24] Подземные виды развили тела, обтекаемые для рытья нор, и в конечном итоге потеряли конечности. [24] Согласно этой гипотезе, такие особенности, как прозрачные , сросшиеся веки ( бриллы ) и потеря наружных ушей, развились, чтобы справиться с трудностями, связанными с окаменением , такими как царапины на роговице и грязь в ушах. [22] [24] Известно, что некоторые примитивные змеи обладали задними конечностями, но их тазовые кости не имели прямого соединения с позвонками. К ним относятся ископаемые виды, такие как Haasiophis , Pachyrhachis и Eupodophis , которые немного старше Наджаша . [19]
Эта гипотеза была подкреплена в 2015 году открытием в Бразилии окаменелости четвероногой змеи возрастом 113 миллионов лет, получившей название Tetrapodophis amplectus . У него много змееподобных черт, он приспособлен к рытью нор, а его желудок указывает на то, что он охотился на других животных. [25] В настоящее время неясно, является ли Tetrapodophis змеей или другим видом из отряда чешуйчатых , поскольку змееподобное тело эволюционировало независимо по крайней мере 26 раз. Tetrapodophis не имеет характерных змеиных черт позвоночника и черепа. [26] [27] Исследование, проведенное в 2021 году, отнесло это животное к группе вымерших морских ящериц мелового периода, известных как долихозавры и не имеющих прямого отношения к змеям. [28]
Альтернативная гипотеза, основанная на морфологии , предполагает, что предки змей были связаны с мозазаврами — вымершими водными рептилиями мелового периода — образующими кладу Pythonomorpha . [18] Согласно этой гипотезе, считается, что сросшиеся прозрачные веки змей эволюционировали для борьбы с морскими условиями (потеря воды роговицей в результате осмоса), а наружные уши были потеряны из-за неиспользования в водной среде. В конечном итоге это привело к появлению животного, похожего на сегодняшних морских змей . В позднем меловом периоде змеи вновь заселили землю и продолжили превращаться в сегодняшних змей. Ископаемые останки змей известны из морских отложений раннего позднего мела, что согласуется с этой гипотезой; тем более, что они старше наземного Наджаша рионегрины . Сходное строение черепа, уменьшенные или отсутствующие конечности и другие анатомические особенности, обнаруженные как у мозазавров, так и у змей, приводят к положительной кладистической корреляции, хотя некоторые из этих особенностей являются общими с варанидами. [ нужна цитата ]
Генетические исследования последних лет показали, что змеи не так тесно связаны с варанами, как когда-то считалось, и, следовательно, не с мозазаврами, предполагаемыми предками в водном сценарии их эволюции. Однако больше доказательств связывают мозазавров со змеями, чем с варанидами. Фрагментированные останки, найденные в юрском и раннем меловом периодах, указывают на более глубокие летописи окаменелостей этих групп, что потенциально может опровергнуть любую гипотезу. [29] [30]
И окаменелости, и филогенетические исследования показывают, что змеи произошли от ящериц , поэтому возник вопрос, какие генетические изменения привели к потере конечностей у предков змей. Потеря конечностей на самом деле очень распространена среди современных рептилий и случалась десятки раз у сцинков , ангвидов и других ящериц. [31]
В 2016 году два исследования показали, что потеря конечностей у змей связана с мутациями ДНК в регуляторной последовательности зоны поляризационной активности (ZRS), регуляторной области гена звукового ежа , который критически необходим для развития конечностей. У более продвинутых змей нет остатков конечностей, но у базальных змей, таких как питоны и удавы, действительно есть следы сильно редуцированных рудиментарных задних конечностей. У эмбрионов питона даже есть полностью развитые зачатки задних конечностей, но их дальнейшее развитие останавливается из-за мутаций ДНК в ZRS. [32] [33] [34] [35]
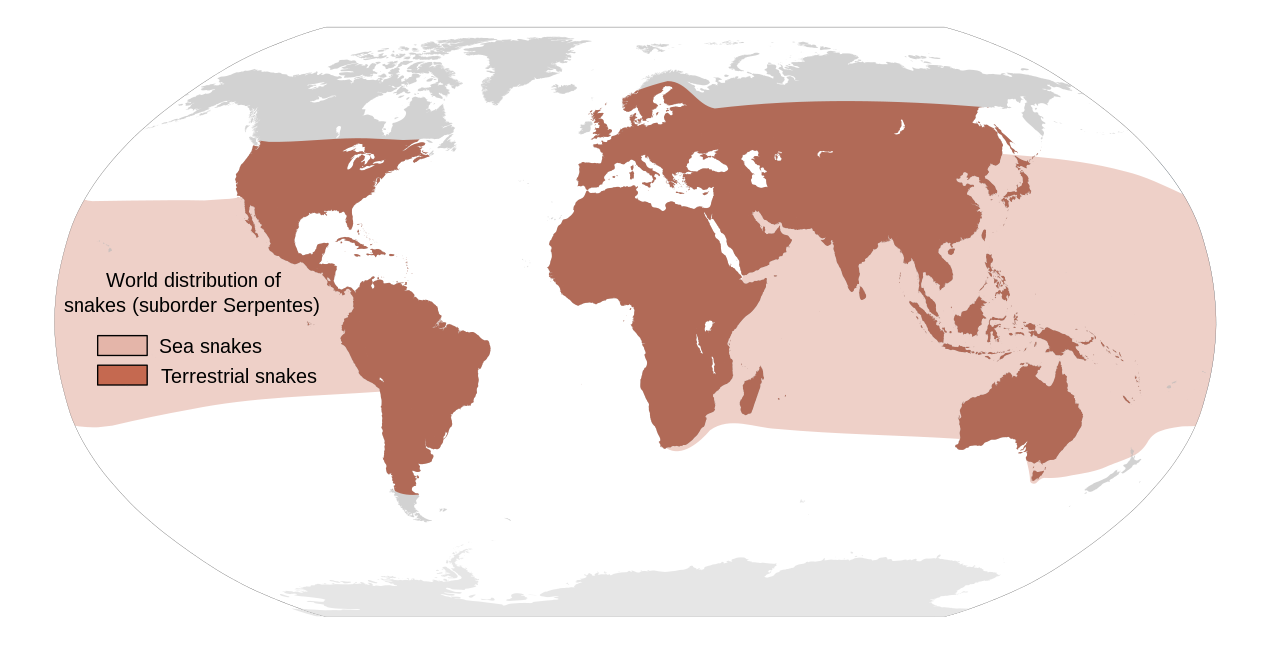
Существует около 3900 видов змей, [36] простирающихся на север до Полярного круга в Скандинавии и на юг через Австралию. [18] Змей можно встретить на всех континентах, кроме Антарктиды, а также в море и на высоте до 16 000 футов (4900 м) в Гималайских горах Азии. [18] [37] : 143 Есть множество островов, на которых змеи отсутствуют, например, Ирландия , Исландия и Новая Зеландия [4] [37] (хотя северные воды Новой Зеландии нечасто посещаются желтобрюхой морской змеей и полосатый морской крайт ). [38]
Все современные змеи сгруппированы в подотряде Serpentes в таксономии Линнея , входящем в отряд Чешуйчатые , хотя их точное размещение внутри чешуйчатых остается спорным. [39]
Два инфраотряда Serpentes — это Alethinophidia и Scolecophidia . [39] Это разделение основано на морфологических характеристиках и сходстве последовательностей митохондриальной ДНК . Алетинофидии иногда разделяют на Henophidia и Caenophidia , причем последние состоят из «колубридных» змей ( колобриды , гадюки , элапиды , гидрофииды и атрактаспиды ) и акрохордид, в то время как другие семейства алетинофидий включают Henophidia. [40] Хотя Madtsoiidae , семейство гигантских, примитивных, похожих на питона змей, не сохранилось сегодня, оно существовало примерно 50 000 лет назад в Австралии и было представлено такими родами, как Wonambi .
Внутри группы ведутся многочисленные дискуссии по систематике. Например, многие источники классифицируют Boidae и Pythonidae как одно семейство, в то время как некоторые по практическим соображениям разделяют Elapidae и Hydrophiidae (морские змеи), несмотря на их чрезвычайно близкое родство.
Недавние молекулярные исследования подтверждают монофилию клад современных змей, сколекофидий, тифлопидов + аномалепидид, алетинофидий, основных алетинофидий, уропельтид ( Cylindrophis , Anomochilus , uropeltines), макростоматанов, буоидов, боидов, питонидов и ценофидий. [14]
Хотя змеи являются рептилиями без конечностей, произошедшими от ящериц (и сгруппированными с ними), существует множество других видов ящериц, которые потеряли свои конечности независимо, но внешне внешне похожи на змей. К ним относятся медленный червь и стеклянная змея .
Другие змеевидные четвероногие , не связанные со змеями, включают червяг (амфибий), земноводных (чешуйчатых, близких к ящерицам) и вымерших аистоподов (амфибий).

Ныне вымерший Titanoboa cerrejonensis имел длину 12,8 м (42 фута). [8] Для сравнения, самыми крупными из ныне живущих змей являются сетчатый питон длиной около 6,95 м (22,8 фута) [7] и зеленая анаконда длиной около 5,21 м (17,1 фута) и считающаяся самой тяжелой змеей на Земле. Земля весом 97,5 кг (215 фунтов). [44]
На другом конце шкалы находится самая маленькая из ныне живущих змей — Leptotyphlops carlae , длина которой составляет около 10,4 см (4,1 дюйма). [6] Большинство змей — довольно маленькие животные, примерно 1 м (3,3 фута) в длину. [45]

Змейские гадюки, питоны и некоторые удавы имеют чувствительные к инфракрасному излучению рецепторы в глубоких бороздках на морде, что позволяет им «видеть» излучаемое тепло теплокровной добычи. У гадюк бороздки расположены между ноздрей и глазом в большой «ямке» с каждой стороны головы. У других змей, чувствительных к инфракрасному излучению, есть несколько небольших губных ямок, выстилающих верхнюю губу, чуть ниже ноздрей. [46]
Змея выслеживает свою добычу с помощью обоняния, собирая частицы в воздухе своим раздвоенным языком , а затем передавая их в сошниково-носовой орган или орган Якобсона во рту для исследования. [46] Вилка языка обеспечивает своего рода направленное обоняние и вкус одновременно. [46] Язык змеи постоянно находится в движении, отбирая частицы из воздуха, земли и воды, анализируя найденные химические вещества и определяя присутствие добычи или хищников в местной среде. У водных змей, таких как анаконда , язык эффективно функционирует под водой. [46]

Нижняя часть змеи очень чувствительна к вибрации, что позволяет ей обнаруживать приближающихся животных, ощущая слабые вибрации земли. [46] Несмотря на отсутствие внешних ушей, они также способны улавливать звуки, передающиеся по воздуху. [47]
Зрение змей сильно различается у разных видов. Некоторые из них обладают острым зрением, а другие способны только отличать свет от темноты, но важной тенденцией является то, что зрительное восприятие змей достаточно адекватно, чтобы отслеживать движения. [48] Как правило, зрение лучше всего у древесных змей и самое слабое у роющих змей. Некоторые обладают бинокулярным зрением , при котором оба глаза способны фокусироваться на одной и той же точке, примером тому является азиатская виноградная змея . Большинство змей фокусируются, перемещая хрусталик вперед и назад относительно сетчатки . У дневных змей зрачки круглые, а у многих ночных змей зрачки щелевые. Большинство видов обладают тремя зрительными пигментами и, вероятно, способны видеть два основных цвета при дневном свете. Кольчатые морские змеи и род Helicops , по-видимому, восстановили большую часть своего цветового зрения в результате адаптации к морской среде, в которой они живут. [49] [50] Был сделан вывод, что последние общие предки всех змей были чувствительны к ультрафиолетовому излучению . зрение, но большинство змей, охота которых зависит от зрения при дневном свете, развили линзы, которые действуют как солнцезащитные очки и фильтруют ультрафиолетовый свет, что, вероятно, также обостряет их зрение за счет улучшения контраста . [51] [52]
Кожа змеи покрыта чешуей . Вопреки популярному представлению о том, что змеи склизкие (из-за того, что змей можно спутать с червями ), змеиная кожа имеет гладкую сухую текстуру. Большинство змей используют для передвижения специальные чешуйки на брюшке, позволяющие им цепляться за поверхности. Чешуя тела может быть гладкой, килевидной или зернистой. Веки змеи представляют собой прозрачные «очковые» чешуйки, также известные как брилле , которые остаются постоянно закрытыми.
Сбрасывание чешуи называется шелушением (или, при обычном использовании, линькой или шелушением ). Змеи сбрасывают весь внешний слой кожи целиком. [53] Змеиные чешуйки не являются отдельными, а являются продолжением эпидермиса — следовательно, они сбрасываются не отдельно, а в виде целого внешнего слоя во время каждой линьки, что похоже на выворачивание носка наизнанку. [54]
Змеи имеют большое разнообразие окраски кожи, которая часто связана с поведением, например, с тенденцией бежать от хищников. Змеи, которые подвергаются высокому риску нападения хищников, обычно имеют однотонную окраску или имеют продольные полосы, что дает мало ориентиров для хищников и позволяет змее уйти незамеченной. Обычные змеи обычно применяют активную стратегию охоты, поскольку их образ жизни позволяет им посылать жертве мало информации о движении. Пятнистые змеи обычно используют стратегии, основанные на засадах, вероятно, потому, что это помогает им сливаться с окружающей средой с объектами неправильной формы, такими как палки или камни. Пятнистый рисунок также может помочь змеям сливаться с окружающей средой. [55]
Форма и количество чешуек на голове, спине и брюхе часто являются характерными и используются в таксономических целях. Чешуи названы в основном в зависимости от их положения на теле. У «продвинутых» ( ценофидийских ) змей широкие брюшные чешуи и ряды спинных чешуек соответствуют позвонкам , что позволяет их подсчитывать без необходимости вскрытия .

Линька (или «шелушение») служит нескольким целям. Он позволяет заменить старую, изношенную кожу и удалить паразитов, таких как клещи и клещи , которые живут в коже. У змей также было замечено, что линька может быть синхронизирована с циклами спаривания. Линька кожи может выделять феромоны и оживлять цвет и рисунок кожи, повышая привлекательность партнеров. [56] Обновление кожи путем линьки предположительно способствует росту у некоторых животных, например, у насекомых, но в случае со змеями это оспаривается. [54] [57]
Линька происходит периодически на протяжении всей жизни змеи. Перед каждой линькой змея перестает есть и часто прячется или перемещается в безопасное место. Незадолго до линьки кожа становится тусклой и сухой, а глаза змеи становятся мутными или голубыми. Внутренняя поверхность старой кожи разжижается, в результате чего она отделяется от новой кожи под ней. Через несколько дней глаза проясняются, и змея «вылезает» из своей старой кожи, которая трескается у змеиной пасти. Змея трется своим телом о грубые поверхности, чтобы помочь сбросить старую кожу. Во многих случаях гипсовая кожа отслаивается назад по телу от головы до хвоста целиком, как будто выдергивают носок наизнанку, обнажая новый, более крупный и яркий слой кожи, образовавшийся под ним. [54] [58]
Молодая змея, которая все еще растет, может сбрасывать кожу до четырех раз в год, а более старая змея может сбрасывать кожу только один или два раза в год. [58] На выброшенной коже сохраняется идеальный отпечаток чешуйчатого рисунка, поэтому обычно можно идентифицировать змею по отлитой коже, если она достаточно неповреждена. [54] Это периодическое обновление привело к тому, что змея стала символом исцеления и медицины , как это изображено на Жезле Асклепия . [59]
Подсчет чешуи иногда можно использовать для определения пола змеи, если этот вид не имеет явно выраженного полового диморфизма . Зонд полностью вводят в клоаку , отмечают точку остановки, затем извлекают и измеряют по субкаудальной чешуе . [60] Подсчет чешуек определяет, является ли змея самцом или самкой, поскольку гемипены самца исследуют на другую глубину (обычно длиннее), чем клоака самки. [60] [ нужны разъяснения ]

Скелет большинства змей состоит исключительно из черепа, подъязычной кости, позвоночного столба и ребер, хотя у хенофидийских змей сохраняются остатки таза и задних конечностей.
Череп состоит из прочного и цельного нейрокраниума , к которому многие другие кости прикреплены лишь слабо, особенно очень подвижные кости челюсти, которые облегчают манипулирование и проглатывание крупной добычи. Левую и правую стороны нижней челюсти соединяет только гибкая связка на передних концах, позволяющая им широко разделяться, а задний конец костей нижней челюсти сочленяется с квадратной костью , обеспечивая дальнейшую подвижность. Нижняя челюсть и квадратная кость могут воспринимать вибрации земли; [61] поскольку стороны нижней челюсти могут двигаться независимо друг от друга, змея, опирающаяся челюстью на поверхность, обладает чувствительным стереослуховым восприятием , используемым для определения положения добычи. Путь челюсть-квадрат- стремечко способен обнаруживать вибрации в масштабе ангстрема , несмотря на отсутствие наружного уха и механизма согласования импедансов , обеспечиваемого косточками других позвоночных, для приема вибраций из воздуха. [62] [63]
Подъязычная кость — небольшая кость, расположенная сзади и вентральнее черепа, в «шейной» области, которая служит местом прикрепления мышц языка змеи, как и у всех других четвероногих .
В позвоночнике насчитывается от 200 до 400 позвонков, а иногда и больше. Каждый из позвонков тела имеет по два ребра, сочленяющихся с ними. Хвостовые позвонки сравнительно немногочисленны (часто менее 20% от общего числа) и лишены ребер. Позвонки имеют выступы, обеспечивающие прочное прикрепление мышц, что позволяет передвигаться без помощи конечностей.
Каудальная автотомия (самоампутация хвоста) — особенность, встречающаяся у некоторых ящериц, — у большинства змей отсутствует. [64] В тех редких случаях, когда у змей она действительно существует, каудальная автотомия является межпозвоночной (что означает разделение соседних позвонков), в отличие от таковой у ящериц, которая является внутрипозвоночной, т.е. разрыв происходит вдоль заранее определенной плоскости перелома, присутствующей на позвонке. [65] [66]
У некоторых змей, особенно удавов и питонов, имеются остатки задних конечностей в виде пары тазовых шпор . Эти небольшие, похожие на когти выступы на каждой стороне клоаки являются внешней частью рудиментарного скелета задней конечности, который включает остатки подвздошной и бедренной костей.
Змеи — полифиодонты , зубы которых постоянно сменяются. [67]

Змеи и другие рептилии, не относящиеся к архозаврам ( крокодилы , динозавры + птицы и их союзники), имеют трехкамерное сердце, которое управляет системой кровообращения через левое и правое предсердия и один желудочек. [68] Внутри желудочек разделен на три взаимосвязанные полости: артериальную полость, легочную полость и венозную полость. [69] Венозная полость получает дезоксигенированную кровь из правого предсердия, а артериальная полость получает насыщенную кислородом кровь из левого предсердия. Под венозной полой полостью находится легочная полость, которая перекачивает кровь в легочный ствол. [70]
Сердце змеи заключено в мешочек, называемый перикардом , расположенный в развилке бронхов . Сердце способно двигаться благодаря отсутствию диафрагмы; эта регулировка защищает сердце от потенциального повреждения, когда крупная проглоченная добыча проходит через пищевод . Селезенка прикрепляется к желчному пузырю и поджелудочной железе и фильтрует кровь. Тимус , расположенный в жировой ткани над сердцем, отвечает за выработку иммунных клеток в крови . Сердечно-сосудистая система змей уникальна наличием почечной портальной системы, в которой кровь из хвоста змеи проходит через почки, прежде чем вернуться в сердце. [71]
Рудиментарное левое легкое часто маленькое, а иногда даже отсутствует, поскольку трубчатые тела змей требуют , чтобы все их органы были длинными и тонкими. [71] У большинства видов функционально только одно легкое. Это легкое содержит васкуляризированную переднюю часть и заднюю часть, которая не участвует в газообмене. [71] Это «мешотчатое легкое» используется в гидростатических целях для регулирования плавучести у некоторых водных змей, и его функция остается неизвестной у наземных видов. [71] Многие парные органы, такие как почки или репродуктивные органы , расположены в организме в шахматном порядке, один расположен впереди другого. [71]
У змей нет лимфатических узлов . [71]

Кобры, гадюки и близкородственные виды используют яд , чтобы обездвижить, ранить или убить свою добычу. Яд представляет собой модифицированную слюну , доставляемую через клыки . [17] [72] : 243 Клыки «продвинутых» ядовитых змей, таких как гадюки и элапиды, полые, что позволяет более эффективно вводить яд, а клыки змей с задними клыками, таких как бумсланг, просто имеют бороздку на задней части. край, чтобы направить яд в рану. Змеиные яды часто специфичны для добычи, и их роль в самозащите второстепенна. [17] [72] : 243
Яд, как и все выделения слюны, является предварительным средством, которое инициирует расщепление пищи на растворимые соединения, способствуя правильному пищеварению. Даже неядовитые укусы змей (как и укусы любого животного) вызывают повреждение тканей. [17] [72] : 209
Некоторые птицы, млекопитающие и другие змеи (например, королевские змеи), охотящиеся на ядовитых змей, выработали устойчивость и даже иммунитет к определенным ядам. [17] : 243 Ядовитые змеи включают три семейства змей и не составляют формальной таксономической классификационной группы.
Разговорный термин «ядовитая змея», как правило , является неверным обозначением змей. Яд вдыхается или проглатывается, тогда как яд, вырабатываемый змеями, впрыскивается жертве через клыки. [73] Однако есть два исключения: Rhabdophis изолирует токсины от жаб, которых он поедает, а затем выделяет их из затылочных желез для отпугивания хищников; а небольшая необычная популяция подвязочных змей в американском штате Орегон сохраняет в печени достаточно токсинов от проглоченных тритонов , чтобы быть эффективно ядовитыми для мелких местных хищников (таких как вороны и лисы ). [74]
Змеиные яды представляют собой сложную смесь белков [72] и хранятся в ядовитых железах на затылке. [74] У всех ядовитых змей эти железы открываются протоками в бороздчатые или полые зубы верхней челюсти. [17] : 243 [73] Белки потенциально могут представлять собой смесь нейротоксинов (поражающих нервную систему), гемотоксинов (поражающих систему кровообращения), цитотоксинов (поражающих непосредственно клетки), бунгаротоксинов (связанных с нейротоксинами, но также непосредственно воздействуют на мышечную ткань) и многие другие токсины, по-разному воздействующие на организм. [73] [72] Почти весь змеиный яд содержит гиалуронидазу , фермент, который обеспечивает быструю диффузию яда. [17] : 243
Ядовитые змеи, использующие гемотоксины, обычно имеют клыки в передней части рта, что облегчает им введение яда в жертву. [72] [73] Некоторые змеи, использующие нейротоксины (например, мангровые змеи ), имеют клыки в задней части рта, загнутые назад. [75] Это затрудняет как использование змеей своего яда, так и учёным, чтобы доить её. [73] Однако элапиды, такие как кобры и крайты, являются протероглифами — у них есть полые клыки, которые нельзя поднять к передней части рта, и они не могут «колоть», как гадюка. Они должны фактически укусить жертву. [17] : 242
Было высказано предположение, что все змеи могут быть в определенной степени ядовитыми, при этом безобидные змеи имеют слабый яд и не имеют клыков. [76] Согласно этой теории, большинство змей, помеченных как «неядовитые», считаются безвредными, поскольку у них либо отсутствует метод доставки яда, либо они не способны доставить достаточно яда, чтобы подвергнуть опасности человека. Теория постулирует, что змеи, возможно, произошли от общего предка ящерицы, который был ядовитым, а также что ядовитые ящерицы, такие как монстр Гила , бисерная ящерица , вараны и ныне вымершие мозазавры , возможно, произошли от этого же общего предка. Они делят эту « ядовитую кладу » с различными другими видами ящеров .
Ядовитые змеи делятся на два таксономических семейства:
Существует третье семейство, включающее опистоглифных (заднезубых) змей (как и большинство других видов змей):
Хотя змеи используют широкий спектр репродуктивных способов, все они используют внутреннее оплодотворение . Это достигается с помощью парных раздвоенных гемипенов , которые в перевернутом виде хранятся в хвосте самца. [77] Гемипены часто имеют бороздки, крючки или шипы и предназначены для захвата стенок клоаки самки . [78] [77] Клитор самки змеи состоит из двух структур, расположенных между клоакой и ароматическими железами. [79]
Большинство видов змей откладывают яйца , которые бросают вскоре после кладки. Однако некоторые виды (например, королевская кобра) строят гнезда и после инкубации остаются рядом с вылупившимися птенцами. [77] Большинство питонов обвивают кладки яиц и остаются с ними до тех пор, пока не вылупятся. [80] Самка питона не оставляет яйца, за исключением того, что время от времени греется на солнце или пьет воду. Она даже будет «дрожать», чтобы выработать тепло для инкубации яиц. [80]
Некоторые виды змей являются яйцеживородящими и сохраняют яйца в своем теле до тех пор, пока они не будут почти готовы вылупиться. [81] [82] Некоторые виды змей, такие как удав и зеленая анаконда, являются полностью живородящими , питая своих детенышей через плаценту , а также желточный мешок ; это весьма необычно для рептилий и обычно встречается у акул-реквиемов или плацентарных млекопитающих . [81] [82] Сохранение яиц и живорождение чаще всего связаны с более холодной средой. [77] [82]

Половой отбор у змей демонстрируется 3000 видами, каждый из которых использует разные тактики поиска партнеров. [83] Ритуальный бой между самцами за самок, с которыми они хотят спариться , включает в себя топпинг - поведение, демонстрируемое большинством гадюк, при котором один самец обвивает вертикально поднятую переднюю часть тела своего противника и заставляет его опускаться вниз. Когда змеи переплетаются, часто происходит укус шеи. [84]
Партеногенез — естественная форма размножения, при которой рост и развитие зародышей происходят без оплодотворения. Agkistrodon contortrix (медноголовый) и Agkistrodon piscivorus (ватноротый) могут размножаться путем факультативного партеногенеза , то есть они способны переключаться с полового способа размножения на бесполый . [85] Наиболее вероятным типом партеногенеза является аутомиксис с терминальным слиянием, процесс, при котором два терминальных продукта одного и того же мейоза сливаются с образованием диплоидной зиготы . Этот процесс приводит к гомозиготности по всему геному , экспрессии вредных рецессивных аллелей и часто к аномалиям развития. Как рожденные в неволе, так и дикие медноголовые и ватные ротики, по-видимому, способны к этой форме партеногенеза. [85]
Размножение у чешуйчатых рептилий почти исключительно половое. Мужчины обычно имеют пару ZZ хромосом, определяющих пол, а женщины - пару ZW. Однако колумбийский радужный удав ( Epicrates maurus ) также может размножаться путем факультативного партеногенеза, что приводит к образованию потомства самок WW. [86] Самки WW, вероятно, производятся терминальным аутомиксисом.

Эмбриональное развитие змеи первоначально следует тем же этапам, что и эмбрионы любого позвоночного . Эмбрион змеи начинается с зиготы , подвергается быстрому делению клеток, образует зародышевый диск , также называемый бластодиском, затем подвергается гаструляции , нейруляции и органогенезу . [88] Деление и пролиферация клеток продолжаются до тех пор, пока не разовьется ранний эмбрион змеи и не будет наблюдаться типичная форма тела змеи. [88] Множество особенностей отличают эмбриологическое развитие змей от других позвоночных, двумя важными факторами являются удлинение тела и отсутствие развития конечностей.

Удлинение тела змей сопровождается значительным увеличением количества позвонков (у мышей их 60, а у змей — более 300). [87] Это увеличение позвонков происходит из-за увеличения сомитов во время эмбриогенеза, что приводит к увеличению количества развивающихся позвонков. [87] Сомиты образуются в пресомитной мезодерме благодаря набору колебательных генов, которые управляют часами сомитогенеза . Часы сомитогенеза змей работают с частотой, в 4 раза превышающей таковую у мыши (после поправки на время развития), создавая больше сомитов и, следовательно, больше позвонков. [87] Считается, что эта разница в тактовой частоте вызвана различиями в экспрессии маргинального гена Lunatic , гена, участвующего в часах сомитогенеза. [89]
Существует обширная литература, посвященная развитию/отсутствию развития конечностей у эмбрионов змей и экспрессии генов, связанной с различными стадиями. У базальных змей , таких как питон, эмбрионы на раннем этапе развития имеют зачаток задних конечностей , который развивается с некоторым количеством хряща и хрящевым тазовым элементом, однако перед вылуплением он дегенерирует. [90] Наличие рудиментарного развития предполагает, что некоторые змеи все еще подвергаются редукции задних конечностей, прежде чем они будут уничтожены. [91] Нет никаких доказательств наличия у базальных змей зачатков передних конечностей и примеров зарождения зачатков передних конечностей змей у эмбриона, поэтому мало что известно о потере этого признака. [91] Недавние исследования показывают, что уменьшение задних конечностей может быть связано с мутациями в энхансерах гена SSH , [91] однако другие исследования показали, что мутации в Hox-генах или их энхансерах могут способствовать потере конечностей у змей. [87] Поскольку многочисленные исследования обнаружили доказательства того, что разные гены играют роль в потере конечностей у змей, вполне вероятно, что множественные мутации генов имели аддитивный эффект, приводящий к потере конечностей у змей [92]

В регионах, где зимы слишком холодные, чтобы змеи могли их терпеть, оставаясь активными, местные виды вступают в период брумации . В отличие от спячки , в которой спящие млекопитающие фактически спят, брумирующие рептилии бодрствуют, но неактивны. Отдельные змеи могут скапливаться в норах, под грудами камней или внутри упавших деревьев, либо большое количество змей может группироваться в гибернакуле .

Все змеи строго плотоядны и охотятся на мелких животных, включая ящериц, лягушек, других змей, мелких млекопитающих, птиц, яйца, рыбу, улиток, червей и насекомых. [17] : 81 [18] [93] Змеи не могут кусать или разрывать пищу на куски, поэтому им приходится проглатывать добычу целиком. На пищевые привычки змеи во многом влияет размер тела; более мелкие змеи едят более мелкую добычу. Молодые питоны могут начать питаться ящерицами или мышами, а во взрослом возрасте превратиться в мелких оленей или антилоп.
Челюсть змеи представляет собой сложную структуру. Вопреки распространенному мнению, что змеи могут вывихивать челюсти, у них чрезвычайно гибкая нижняя челюсть , две половины которой не соединены жестко, и множество других суставов черепа, которые позволяют змее открывать пасть достаточно широко, чтобы глотать. добычу целиком, даже если она в диаметре больше самой змеи. [93] Например, африканская змея, питающаяся яйцами, имеет гибкие челюсти, приспособленные для поедания яиц, размер которых намного превышает диаметр ее головы. [17] : 81 У этой змеи нет зубов, но есть костные выступы на внутреннем крае позвоночника , которые она использует, чтобы разбить скорлупу при поедании яиц. [17] : 81

Большинство змей питаются разнообразными животными, но у определенных видов существует некоторая специализация. Королевские кобры и австралийский хоккей с мячом поедают других змей. У видов семейства Pareidae больше зубов на правой стороне рта, чем на левой, поскольку они в основном охотятся на улиток, а раковины обычно вращаются по часовой стрелке. [17] : 184 [94] [95]
У некоторых змей есть ядовитый укус, которым они убивают добычу перед тем, как ее съесть. [93] [96] Другие змеи убивают свою добычу сужением , [ 93] а некоторые заглатывают добычу, когда она еще жива. [17] : 81 [93]

После еды змеи впадают в спячку, позволяя начать процесс пищеварения ; [60] это интенсивная деятельность, особенно после поедания крупной добычи. У видов, которые питаются лишь спорадически, весь кишечник между приемами пищи переходит в сокращенное состояние для сохранения энергии. Затем пищеварительная система «настраивается» на полную мощность в течение 48 часов после потребления добычи. Будучи экзотермическими («хладнокровными»), температура окружающей среды играет важную роль в процессе пищеварения. Идеальная температура для переваривания пищи змеями — 30 °C (86 °F). В пищеварении у змей задействовано огромное количество метаболической энергии, например, температура поверхности тела южноамериканской гремучей змеи ( Crotalus durissus ) увеличивается на целых 1,2 ° C (2,2 ° F) во время процесса пищеварения. [97] Если змея потревожена после того, как недавно поела, она часто срыгивает добычу, чтобы избежать предполагаемой угрозы. Когда процесс пищеварения не нарушен, он очень эффективен; пищеварительные ферменты змеи растворяют и поглощают все, кроме волос (или перьев) и когтей жертвы, которые выводятся из организма вместе с отходами .
Капюшон (расширение области шеи) является визуальным сдерживающим фактором, чаще всего наблюдаемым у кобр (елапид), и в первую очередь контролируется реберными мышцами. [98] Надевание капюшона может сопровождаться плевком яда в сторону угрожающего объекта, [99] и изданием специального звука; шипение. Исследования кобр в неволе показали, что во время надевания капюшона приподнимается от 13 до 22% длины тела. [100]
Отсутствие конечностей не препятствует передвижению змей. Они разработали несколько различных способов передвижения для работы в определенных условиях. В отличие от походки конечностей животных, которые образуют континуум, каждый способ передвижения змей дискретен и отличается от других; переходы между режимами резкие. [101] [102]

Боковая волнистость — единственный способ передвижения в воде и наиболее распространенный способ передвижения на суше. [102] В этом режиме тело змеи поочередно сгибается влево и вправо, что приводит к серии движущихся назад «волн». [101] Хотя это движение кажется быстрым, редко регистрируется, что змеи двигаются быстрее, чем на две длины тела в секунду, а часто и намного меньше. [103] Этот способ передвижения имеет те же чистые затраты на транспортировку (сожженные калории на метр перемещения), что и бег ящериц той же массы. [104]
Наземные боковые волнистости - наиболее распространенный способ наземного передвижения большинства видов змей. [101] В этом режиме движущиеся назад волны давят на точки контакта с окружающей средой, такие как камни, ветки, неровности почвы и т. д. [101] Каждый из этих объектов окружающей среды, в свою очередь, генерирует силу реакции, направленную вперед. и к средней линии змеи, что приводит к толчку вперед, в то время как боковые компоненты уравновешиваются. [105] Скорость этого движения зависит от плотности точек толчка в окружающей среде, при этом средняя плотность около 8 [ необходимы пояснения ] по длине змеи является идеальной. [103] Скорость волны точно такая же, как и скорость змеи, и в результате каждая точка на теле змеи повторяет путь точки перед ней, что позволяет змеям перемещаться через очень густую растительность и небольшие отверстия. [105]
Во время плавания волны становятся больше по мере движения вниз по телу змеи, и волна движется назад быстрее, чем змея движется вперед. [106] Тяга создается прижатием тела к воде, что приводит к наблюдаемому скольжению. Несмотря на общее сходство, исследования показывают, что характер мышечной активации различен при водных и наземных боковых волнах, что дает основание называть их отдельными режимами. [107] Все змеи могут двигаться вперед (с волнами, движущимися назад), но наблюдалось только морские змеи, меняющие направление движения (движущиеся назад с волнами, движущимися вперед). [101]

Чаще всего используется колубридными змеями ( колобридами , элапидами и гадюками ), когда змее приходится двигаться в среде, где нет неровностей, на которые можно было бы наталкиваться (что делает невозможными боковые волнистости), например, на скользкой илистой равнине или песчаной дюне. модифицированная форма боковой волнистости, при которой все сегменты тела, ориентированные в одном направлении, остаются в контакте с землей, в то время как другие сегменты поднимаются вверх, что приводит к своеобразному «перекатывающему» движению. [108] [109] Этот способ передвижения позволяет преодолеть скользкость песка или грязи, отталкиваясь только статическими частями тела, тем самым сводя к минимуму скольжение. [108] Статичность точек контакта можно продемонстрировать по следам извивающейся змеи, на которых виден отпечаток каждой чешуи на брюхе, без какого-либо размытия. Этот способ передвижения имеет очень низкую калорийность: менее 1/3 затрат ящерицы на перемещение на то же расстояние . [104] Вопреки распространенному мнению, нет никаких доказательств того, что боковой ветер связан с горячим песком. [108]
Когда точки толчка отсутствуют, но недостаточно места для использования бокового движения из-за боковых ограничений, например, в туннелях, змеи полагаются на движение гармошкой. [101] [109] В этом режиме змея прижимает заднюю часть своего тела к стене туннеля, в то время как передняя часть змеи вытягивается и выпрямляется. [108] Затем передняя часть сгибается и образует точку крепления, а задняя часть выпрямляется и тянется вперед. Этот способ передвижения медленный и очень трудоемкий: затраты на боковые колебания на том же расстоянии в семь раз превышают затраты. [104] Такая высокая стоимость обусловлена повторяющимися остановками и стартами частей тела, а также необходимостью использования активных мышечных усилий для упирания в стены туннеля.

Передвижение змей в древесных местообитаниях изучается лишь недавно. [110] Находясь на ветвях деревьев, змеи используют несколько способов передвижения в зависимости от вида и текстуры коры. [110] В общем, змеи будут использовать модифицированную форму передвижения гармошкой на гладких ветвях, но будут совершать боковые волнистые движения, если доступны точки контакта. [110] Змеи движутся быстрее на маленьких ветвях и при наличии точек контакта, в отличие от животных с конечностями, которые лучше себя чувствуют на больших ветвях с небольшим «беспорядком». [110]
Планирующие змеи ( Chrysopelea ) Юго-Восточной Азии взлетают с кончиков ветвей, расправляя ребра и совершая волнообразные движения в стороны, скользя между деревьями. [108] [111] [112] Эти змеи могут выполнять контролируемое планирование на сотни футов в зависимости от высоты запуска и даже поворачиваться в воздухе. [108] [111]
Самый медленный способ передвижения змеи - это прямолинейное передвижение, которое также является единственным, при котором змее не нужно сгибать свое тело в стороны, хотя она может делать это при повороте. [113] В этом режиме чешуя живота поднимается и вытягивается вперед, а затем опускается вниз, и тело натягивается на нее. Волны движения и стаза проходят назад, вызывая серию пульсаций на коже. [113] Ребра змеи не движутся при этом способе передвижения, и этот метод чаще всего используется крупными питонами , удавами и гадюками при преследовании добычи на открытой местности, поскольку движения змеи тонкие и их труднее обнаружить добыче. таким образом. [108]


Змеи обычно не охотятся на людей. Если они не испуганы и не ранены, большинство змей предпочитают избегать контакта и не нападают на людей. Неядовитые змеи, за исключением крупных удавов, не представляют угрозы для человека. Укус неядовитой змеи обычно безвреден; их зубы не приспособлены для разрывания или нанесения глубокой колотой раны, а скорее для хватания и удержания. Хотя при укусе неядовитой змеи существует возможность заражения и повреждения тканей, ядовитые змеи представляют гораздо большую опасность для человека. [17] : 209 Всемирная организация здравоохранения (ВОЗ) относит укус змеи к категории «других забытых состояний». [116]
Документально подтвержденные случаи смерти в результате укусов змей редки. Несмертельные укусы ядовитых змей могут привести к необходимости ампутации конечности или ее части. Из примерно 725 видов ядовитых змей во всем мире только 250 способны убить человека одним укусом. В Австралии в среднем случается только один смертельный укус змеи в год. В Индии за один год регистрируется 250 000 укусов змей, при этом первоначально зарегистрировано до 50 000 случаев смерти. [117] По оценкам ВОЗ, около 100 000 человек ежегодно умирают в результате укусов змей, и ежегодно укусы змей вызывают примерно в три раза больше ампутаций и других необратимых инвалидностей. [118]
Лечение укуса змеи так же разнообразно, как и сам укус. Самый распространенный и эффективный метод — использование противоядия (или противоядия), сыворотки, приготовленной из яда змеи. Некоторые противоядия являются видоспецифичными (моновалентными), тогда как некоторые предназначены для использования с несколькими видами (поливалентные). В Соединенных Штатах, например, все виды ядовитых змей являются гадюками , за исключением коралловой змеи . Для производства противоядия смесь ядов различных видов гремучих змей , медноголовых и хлопчатников вводят в тело лошади во все возрастающих дозах до тех пор, пока лошадь не будет иммунизирована. Затем у иммунизированной лошади берут кровь. Сыворотку отделяют, дополнительно очищают и лиофилизируют. Он восстанавливается стерильной водой и становится противоядием. По этой причине люди, страдающие аллергией на лошадей, с большей вероятностью будут иметь аллергическую реакцию на противоядие. [119] Противоядие для более опасных видов (таких как мамбы , тайпаны и кобры ) производится аналогичным образом в Южной Африке, Австралии и Индии, хотя эти противоядия являются видоспецифичными.

В некоторых частях света, особенно в Индии, заклинание змей представляет собой придорожное представление, проводимое заклинателем. В таком представлении заклинатель змей несет корзину со змеей, которую он, по-видимому, очаровывает, играя мелодии на своем музыкальном инструменте, похожем на флейту, на что змея реагирует. [120] На самом деле змея реагирует на движение флейты, а не на звук, который она издает, поскольку у змей нет внешних ушей (хотя у них есть внутренние уши). [120]
Закон о защите дикой природы 1972 года в Индии технически запрещает заклинание змей по причине уменьшения жестокого обращения с животными. Другие типы заклинателей змей используют шоу змей и мангустов , где два животных инсценируют драку; однако это случается не очень часто, поскольку животные могут быть серьезно ранены или убиты. Профессия заклинания змей вымирает в Индии из-за конкуренции со стороны современных форм развлечений и законов об окружающей среде, запрещающих эту практику. Многие индийцы никогда не видели очаровательных змей, и это становится сказкой прошлого. [120] [121] [122] [123]
Племя ирулас из Андхра-Прадеша и Тамилнада в Индии занималось охотниками-собирателями в жарких, сухих равнинных лесах и на протяжении поколений практиковало искусство ловли змей. У них есть обширные знания о змеях в этой области. Обычно они ловят змей с помощью простой палки. Ранее Ирулас вылавливали тысячи змей для производства змеиной кожи. После полного запрета производства змеиной кожи в Индии и защиты всех змей в соответствии с Законом о защите дикой природы Индии 1972 года они сформировали Кооператив ловцов змей Ирула и переключились на ловлю змей для удаления яда, выпуская их в дикую природу после четыре извлечения. Собранный таким образом яд используется для производства спасительного противоядия, биомедицинских исследований и других лекарственных средств. [124] Также известно, что ирулы едят некоторых змей, которых ловят, и очень полезны при истреблении крыс в деревнях .
Несмотря на существование заклинателей змей, были также профессиональные ловцы змей или дрессировщики . В современном отлове змей герпетолог использует длинную палку с V-образным концом. Некоторые ведущие телешоу, такие как Билл Хааст , Остин Стивенс , Стив Ирвин и Джефф Корвин , предпочитают ловить их голыми руками.

Хотя змей обычно не считают пищей, в некоторых культурах их употребление приемлемо и даже может считаться деликатесом. Змеиный суп популярен в кантонской кухне , его едят местные жители осенью, чтобы согреть тело. В западных культурах зафиксировано употребление змей только в крайних случаях голода, [125] за исключением приготовленного мяса гремучей змеи , которое обычно употребляют в пищу в Техасе [126] и некоторых частях Среднего Запада США .

В азиатских странах, таких как Китай, Тайвань, Таиланд, Индонезия, Вьетнам и Камбоджа, считается, что употребление крови змей, особенно кобры, повышает сексуальную потенцию. [127] Если это возможно, кровь сливают, пока кобра еще жива, и обычно ее смешивают с каким-либо ликером для улучшения вкуса. [127]
Использование змей в алкоголе принято в некоторых странах Азии. В таких случаях одну или несколько змей оставляют настаиваться в банке или контейнере с ликером, поскольку, как утверждается, это делает ликер более крепким (а также более дорогим). Одним из примеров этого является змея Хабу , которую иногда добавляют в окинавский ликер Хабусю (ハブ酒), также известный как «Хабу Сакэ». [128]
Змеиное вино (蛇酒) — алкогольный напиток, производимый путем настаивания целых змей в рисовом вине или зерновом спирте . Впервые было зарегистрировано, что его употребляли в Китае во времена династии Западная Чжоу . Этот напиток считается важным лечебным средством и, согласно традиционной китайской медицине, восстанавливает силы человека . [129]
В западном мире некоторых змей держат в качестве домашних животных, особенно послушных видов, таких как шаровидный питон и кукурузная змея . Чтобы удовлетворить спрос, была развита индустрия разведения в неволе . Змеи, выращенные в неволе, считаются более предпочтительными, чем особи, пойманные в дикой природе, и, как правило, из них получаются лучшие домашние животные. [130] По сравнению с более традиционными видами домашних животных, змеи могут быть домашними животными, не требующими особого ухода; им требуется минимальное пространство, поскольку длина большинства распространенных видов не превышает 5 футов (1,5 м), и их можно кормить относительно нечасто - обычно один раз в пять-14 дней. Продолжительность жизни некоторых змей при правильном уходе превышает 40 лет.



.svg/1280px-Brazilian_Expeditionary_Forces_insignia_(smoking_snake).svg.png)

В древней Месопотамии Нира , бог-посланник Иштарана , изображался в виде змеи на кудуррусе , или пограничных камнях . [131] Изображения двух переплетенных змей распространены в шумерском и неошумерском искусстве [131] и до сих пор время от времени появляются на цилиндрических печатях и амулетах вплоть до тринадцатого века до нашей эры. [131] Рогатая гадюка ( Cerastes cerastes ) появляется в касситском и неоассирийском кудуррусе [131] и упоминается в ассирийских текстах как магическое защитное существо. [131] Драконоподобное существо с рогами, телом и шеей змеи, передними лапами льва и задними лапами птицы появляется в месопотамском искусстве от аккадского периода до эллинистического периода ( 323 г. до н.э. – 31 г.). ДО Н.Э). [131] Это существо, известное на аккадском языке как mušḫuššu , что означает «яростный змей», использовалось как символ определенных божеств, а также как общая защитная эмблема. [131] Первоначально он, по-видимому, был помощником бога Подземного мира Ниназу , [131] но позже стал помощником хурритского бога бури Тишпака , а также, позже, сына Ниназу Нингишзиды , вавилонского национального бога Мардука , бог-писец Набу и ассирийский национальный бог Ашур. [131]
В египетской истории змея занимает первостепенную роль, наряду с нильской коброй, украшавшей в древности корону фараона. Ему поклонялись как одному из богов, а также использовали в зловещих целях: убийство противника и ритуальное самоубийство ( Клеопатра ). [ нужна цитация ] Уроборос был хорошо известным древнеегипетским символом змеи, глотающей свой хвост . [132] Предшественником уробороса был «Многоликий», [132] змей с пятью головами, который, согласно Амдуату , древнейшей сохранившейся Книге загробной жизни , как говорят, обвивался вокруг трупа солнца. бог Ра покровительствует. [132] Самое раннее сохранившееся изображение «истинного» уробороса происходит из позолоченных святынь в гробнице Тутанхамона . [132] В первые века нашей эры уроборос был принят христианами -гностиками в качестве символа, [132] и в главе 136 «Пистис Софии» , раннего гностического текста, описывается «великий дракон, чей хвост находится во рту». [132] В средневековой алхимии уроборос стал типичным западным драконом с крыльями, ногами и хвостом. [132]
В Библии аммонитянский царь Нахаш , имя которого означает «Змей», изображен весьма негативно, как особо жестокий и презренный враг древних евреев.

Древние греки использовали Горгонейон , изображение отвратительного лица со змеями вместо волос, как апотропный символ для отражения зла. [133] В греческом мифе , описанном Псевдо-Аполлодором в его «Библиотеке» , Медуза была Горгоной со змеями вместо волос, чей взгляд обращал всех, кто смотрел на нее, в камень и был убит героем Персеем . [134] [135] [136] В «Метаморфозах» римского поэта Овидия говорится , что Медуза когда-то была прекрасной жрицей Афины , которую Афина превратила в змееволосого монстра после того, как она была изнасилована богом Посейдоном в Афине. храм. [137] В другом мифе, на который ссылается беотийский поэт Гесиод и подробно описанный Псевдо-Аполлодором, говорится, что герой Геракл убил Лернейскую гидру , [138] [139] многоглавую змею, обитавшую в болотах Лерна . [138] [139]
В легендарном рассказе об основании Фив упоминается чудовищная змея, охраняющая источник, из которого новое поселение должно было черпать воду. В битве со змеей и ее убийстве все спутники основателя Кадма погибли, что привело к появлению термина « Кадмейская победа » (то есть победа, влекущая за собой собственную гибель). [ нужна цитата ]
Три медицинских символа со змеями, которые используются до сих пор, — это Чаша Гигеи , символизирующая аптеку, а также Кадуцей и Жезл Асклепия , которые являются символами, обозначающими медицину в целом. [59]
Одна из этимологий, предложенных для общего женского имени Линда, заключается в том, что оно может происходить от древнегерманского Lindi или Linda , что означает змея.
Индию часто называют страной змей, и она богата традициями в отношении змей. [140] Змеям даже сегодня поклоняются как богам, и многие женщины выливают молоко в змеиные ямы (несмотря на отвращение змей к молоку). [140] Кобру можно увидеть на шее Шивы , а Вишну часто изображают спящим на семиглавой змее или в кольцах змеи. [141] В Индии также есть несколько храмов исключительно для кобр, которых иногда называют Наградж (Король змей), и считается, что змеи являются символами плодородия. Ежегодно проводится индуистский фестиваль Наг Панчами , в этот день почитают змей и молятся им. См. также Нага . [ нужна цитата ]
В Индии существует еще одна мифология о змеях. На хинди широко известны как змеи « Иччхадхари ». Такие змеи могут принимать облик любого живого существа, но предпочитают человеческий облик. Эти мифические змеи обладают ценным драгоценным камнем под названием «Мани», который по блеску превосходит алмаз. В Индии ходит много историй о жадных людях, которые пытались завладеть этим драгоценным камнем и в конечном итоге были убиты. [ нужна цитата ]
Змея – одно из 12 небесных животных китайского зодиака в китайском календаре . [142]
Многие древние перуанские культуры поклонялись природе. [143] В своем искусстве они подчеркивали значение животных и часто изображали змей. [144]
Змеи используются в индуизме как часть ритуального поклонения. [145] На ежегодном фестивале Наг Панчами участники поклоняются либо живым кобрам, либо изображениям Нагов . Господь Шива на большинстве изображений изображен со змеей, обвивающей его шею. [146] Пураническая литература включает в себя различные истории, связанные со змеями, например, говорят, что Шеша держит все планеты Вселенной на своих капюшонах и постоянно воспевает славу Вишну изо всех своих уст. Другими известными змеями в индуизме являются Васуки , Такшака , Каркотака и Пингала . Термин Нага используется для обозначения существ, принимающих форму больших змей в индуизме и буддизме . [147]
Змеи широко почитались во многих культурах, например, в Древней Греции , где змей считался целителем. [148] Асклепий носил вокруг своей палочки змеиную рану, символ, который сегодня можно увидеть на многих машинах скорой помощи. [149] В иудаизме медная змея также является символом исцеления, спасения жизни от неминуемой смерти. [150]
С религиозной точки зрения змея и ягуар были, возможно, самыми важными животными в древней Мезоамерике . [151] «В состояниях экстаза лорды танцуют змеиный танец; огромные нисходящие змеи украшают и поддерживают здания от Чичен-Ицы до Теночтитлана , а слово коатль на языке науатль , означающее змею или близнеца, является частью первичных божеств, таких как Мишкоатль , Кетцалькоатль и Коатликуэ ». [152] В календарях майя и ацтеков пятый день недели был известен как День Змеи.
В некоторых частях христианства искупительная работа Иисуса Христа сравнивается со спасением жизни через созерцание Нехуштана ( медного змея). [153] Обработчики змей используют змей как неотъемлемую часть церковного богослужения, чтобы продемонстрировать свою веру в божественную защиту. Однако чаще всего в христианстве змей изображался как представитель зла и коварных заговоров, как это видно из описания в книге Бытия змея, искушающего Еву в Эдемском саду . [154] Предполагается, что Святой Патрик изгнал всех змей из Ирландии во время обращения страны в христианство в V веке, что объясняет отсутствие там змей. [155]
В христианстве и иудаизме змея появляется в первой книге Библии, когда змей появляется перед Адамом и Евой и искушает их запретным плодом с Древа познания . [154] Змея возвращается в Книге Исход , когда Моисей превращает свой посох в змею в знак Божьей силы, а позже, когда он делает Нехуштан, бронзовую змею на шесте, которая, глядя на нее, исцеляла людей от укусов змеи, преследовавшие их в пустыне. Змей в последний раз появляется, символизируя сатану, в Книге Откровения : «И схватил дракона, старого змея, который есть дьявол и сатана, и связал его на тысячу лет». [156]
В неоязычестве и Викке змея рассматривается как символ мудрости и знаний. [157] Кроме того, змей иногда связывают с Гекатой , греческой богиней колдовства . [158]

Несколько соединений змеиных ядов исследуются в качестве потенциальных средств лечения или профилактики боли, рака, артрита, инсульта, болезней сердца, гемофилии и гипертонии, а также для контроля кровотечений (например, во время операций). [160] [161] [162]




{kind=link}
{kind=link}