Эмбриональное развитие человека или эмбриогенез человека — это развитие и формирование человеческого эмбриона . Оно характеризуется процессами деления клеток и клеточной дифференциации эмбриона, которые происходят на ранних стадиях развития. С биологической точки зрения развитие человеческого организма влечет за собой рост от одноклеточной зиготы до взрослого человека . Оплодотворение происходит, когда сперматозоид успешно проникает в яйцеклетку (яйцеклетку) и сливается с ней . Затем генетический материал сперматозоида и яйцеклетки объединяется, образуя одноклеточную зиготу, и начинается зародышевая стадия развития. Эмбриональное развитие человека охватывает первые восемь недель развития, которые имеют 23 стадии, называемые стадиями Карнеги . В начале девятой недели эмбрион называется плодом ( в британском английском пишется «foetus» ). По сравнению с эмбрионом плод имеет более узнаваемые внешние черты и более полный набор развивающихся органов.
Эмбриология человека изучает это развитие в течение первых восьми недель после оплодотворения. Нормальный период гестации (беременности) составляет около девяти месяцев или 40 недель.
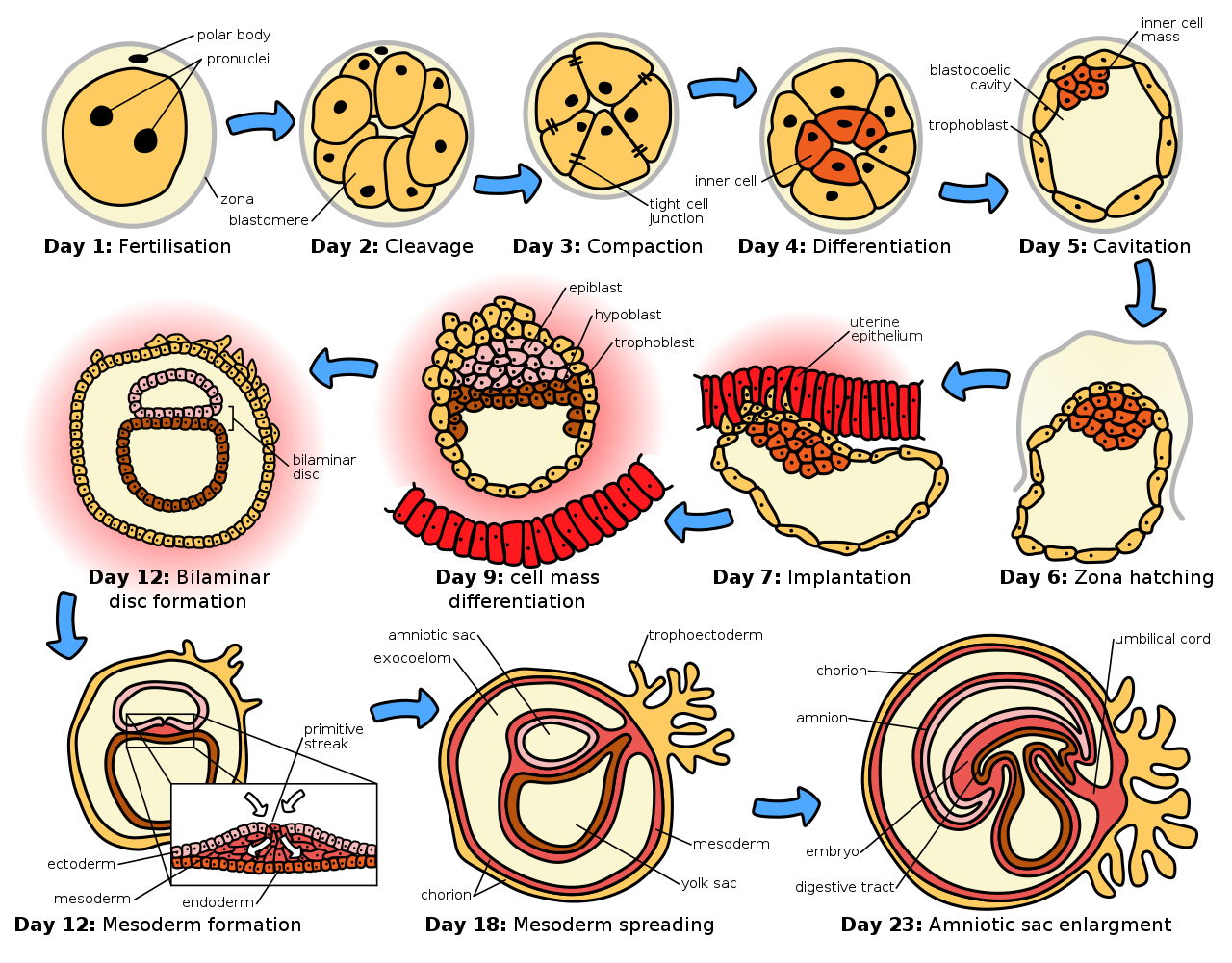
Зародышевая стадия относится ко времени от оплодотворения через развитие раннего эмбриона до завершения имплантации в матку . Зародышевая стадия занимает около 10 дней. [1] На этой стадии зигота делится в процессе, называемом дроблением . Затем формируется бластоциста , которая имплантируется в матку . Эмбриогенез продолжается следующей стадией гаструляции , когда три зародышевых слоя эмбриона формируются в процессе, называемом гистогенезом , и следуют процессы нейруляции и органогенеза .
Весь процесс эмбриогенеза включает в себя скоординированные пространственные и временные изменения в экспрессии генов , росте клеток и клеточной дифференциации . Почти идентичный процесс происходит у других видов, особенно среди хордовых .
Оплодотворение происходит, когда сперматозоид успешно проникает в яйцеклетку и два набора генетического материала, переносимые гаметами, сливаются вместе, в результате чего образуется зигота (одна диплоидная клетка). Обычно это происходит в ампуле одной из маточных труб . Зигота содержит объединенный генетический материал, переносимый как мужскими, так и женскими гаметами, который состоит из 23 хромосом из ядра яйцеклетки и 23 хромосом из ядра сперматозоида. 46 хромосом претерпевают изменения перед митотическим делением , которое приводит к образованию эмбриона, имеющего две клетки.
Успешное оплодотворение обеспечивается тремя процессами, которые также действуют как контроль для обеспечения видоспецифичности. Первый — это хемотаксис , который направляет движение сперматозоида к яйцеклетке. [2] Во-вторых, происходит адгезионная совместимость между сперматозоидом и яйцеклеткой. Когда сперматозоид прилипает к яйцеклетке, происходит третий процесс акросомной реакции ; передняя часть головки сперматозоида покрывается акросомой , которая содержит пищеварительные ферменты , разрушающие zona pellucida и обеспечивающие его проникновение. [3] Проникновение сперматозоида вызывает высвобождение кальция, который блокирует проникновение других сперматозоидов. [3] Параллельная реакция происходит в яйцеклетке, называемая zona reaction . Это приводит к высвобождению кортикальных гранул , которые выделяют ферменты, переваривающие белки рецепторов сперматозоидов, тем самым предотвращая полиспермию . [4] Гранулы также сливаются с плазматической мембраной и модифицируют zona pellucida таким образом, чтобы предотвратить дальнейшее проникновение сперматозоидов.

Начало процесса дробления отмечается, когда зигота делится посредством митоза на две клетки. Этот митоз продолжается, и первые две клетки делятся на четыре клетки, затем на восемь клеток и так далее. Каждое деление занимает от 12 до 24 часов. Зигота крупнее любой другой клетки и подвергается дроблению без какого-либо общего увеличения размера. Это означает, что с каждым последующим подразделением соотношение ядерного и цитоплазматического материала увеличивается. [5]
Первоначально делящиеся клетки, называемые бластомерами ( по-гречески blastos — росток), недифференцированы и объединены в сферу, заключенную в zona pellucida яйцеклетки. Когда сформировано восемь бластомеров , они начинают уплотняться . [6] Они начинают развивать щелевые соединения , что позволяет им развиваться интегрированным образом и координировать свою реакцию на физиологические сигналы и сигналы окружающей среды. [7]
Когда количество клеток составляет около шестнадцати, твердая сфера клеток внутри блестящей оболочки называется морулой . [ 8]

Дробление само по себе является первой стадией бластуляции , процесса формирования бластоцисты . Клетки дифференцируются во внешний слой клеток, называемый трофобластом , и внутреннюю клеточную массу . При дальнейшем уплотнении отдельные внешние бластомеры, трофобласты, становятся неразличимыми. Они по-прежнему заключены в zona pellucida . Это уплотнение служит для того, чтобы сделать структуру водонепроницаемой, содержащей жидкость, которую клетки позже будут выделять. Внутренняя масса клеток дифференцируется, чтобы стать эмбриобластами и поляризуется на одном конце. Они смыкаются и образуют щелевые контакты , которые облегчают клеточную коммуникацию. Эта поляризация оставляет полость, бластоцель , создавая структуру, которая теперь называется бластоцистой. (У животных, кроме млекопитающих, это называется бластула ) .
Трофобласты выделяют жидкость в бластоцель. В результате этого бластоциста увеличивается в размерах, что приводит к ее вылуплению через zona pellucida, которая затем распадается. [5] Этот процесс называется zona hatching и происходит на шестой день развития эмбриона, непосредственно перед процессом имплантации. Вылупление человеческого эмбриона поддерживается протеазами, выделяемыми клетками бластоцисты, которые переваривают белки zona pellucida, в результате чего образуется отверстие. Затем из-за ритмичного расширения и сокращения бластоцисты происходит повышение давления внутри самой бластоцисты, отверстие расширяется, и, наконец, бластоциста может выйти из этой жесткой оболочки.
Внутренняя клеточная масса даст начало преэмбриону , [ 9] амниону , желточному мешку и аллантоису , в то время как плодная часть плаценты сформируется из внешнего слоя трофобласта. Эмбрион вместе с его оболочками называется зародышем , и на этой стадии зародыш достигает матки . В конечном итоге блестящая оболочка полностью исчезает, и теперь открытые клетки трофобласта позволяют бластоцисте прикрепиться к эндометрию , где она имплантируется . Формирование гипобласта и эпибласта , которые являются двумя основными слоями двухслойного зародышевого диска, происходит в начале второй недели. [10] И эмбриобласт, и трофобласт превратятся в два подслоя. [11] Внутренние клетки превратятся в слой гипобласта, который окружит другой слой, называемый эпибластом, и эти слои сформируют зародышевый диск, который разовьется в эмбрион. [10] [11]
Трофобласт также разовьет два подслоя: цитотрофобласт , который находится перед синцитиотрофобластом , который, в свою очередь, находится внутри эндометрия . [10] Затем появится еще один слой, называемый экзоцеломической мембраной или мембраной Гейзера , который окружит цитотрофобласт, а также примитивный желточный мешок. [11] Синцитиотрофобласт будет расти и войдет в фазу, называемую лакунарной стадией, в которой появятся некоторые вакуоли, которые в последующие дни заполнятся кровью. [10] [11] Развитие желточного мешка начинается с гипобластических плоских клеток, которые образуют экзоцеломическую мембрану, которая покроет внутреннюю часть цитотрофобласта, образуя примитивный желточный мешок. Эрозия эндотелиальной выстилки материнских капилляров синцитиотрофобластическими клетками приводит к образованию материнских синусоидов, откуда кровь начинает проникать и течь в трофобластические лакуны и через них, давая начало маточно-плацентарному кровообращению. [12] [13] Впоследствии новые клетки, полученные из желточного мешка, будут установлены между трофобластом и экзоцеломической мембраной и дадут начало внеэмбриональной мезодерме , которая сформирует хорионическую полость . [11]
В конце второй недели развития некоторые клетки трофобласта проникают и образуют округлые столбики в синцитиотрофобласте. Эти столбики известны как первичные ворсинки . В то же время другие мигрирующие клетки формируют в экзоцеломической полости новую полость, называемую вторичным или окончательным желточным мешком, меньшим, чем примитивный желточный мешок. [11] [12]

После овуляции эндометриальный слой трансформируется в секреторный слой, готовясь принять эмбрион. Он утолщается, его секреторные железы удлиняются и становятся все более васкуляризированными . Этот слой полости матки (или матки) теперь известен как децидуальная оболочка , и он производит большое количество крупных децидуальных клеток в своей увеличенной межжелезистой ткани. Бластомерные клетки в бластоцисте расположены во внешнем слое, называемом трофобластом . Затем трофобласт дифференцируется во внутренний слой, цитотрофобласт , и внешний слой, синцитиотрофобласт . Цитотрофобласт содержит кубовидные эпителиальные клетки и является источником делящихся клеток , а синцитиотрофобласт представляет собой синцитиальный слой без клеточных границ.
Синцитиотрофобласт имплантирует бластоцисту в децидуальный эпителий с помощью проекций хорионических ворсин , образуя эмбриональную часть плаценты. Плацента развивается после имплантации бластоцисты, соединяя эмбрион со стенкой матки. Децидуальная оболочка здесь называется decidua basalis; она находится между бластоцистой и миометрием и образует материнскую часть плаценты . Имплантация осуществляется с помощью гидролитических ферментов , которые разрушают эпителий . Синцитиотрофобласт также вырабатывает хорионический гонадотропин человека , гормон , который стимулирует высвобождение прогестерона из желтого тела . Прогестерон обогащает матку толстой оболочкой кровеносных сосудов и капилляров, чтобы она могла насыщать кислородом и поддерживать развивающийся эмбрион. Матка высвобождает сахар из запасенного гликогена из своих клеток для питания эмбриона . [14] Ворсинки начинают разветвляться и содержать кровеносные сосуды эмбриона. Другие ворсинки, называемые терминальными или свободными ворсинками, обмениваются питательными веществами. Эмбрион соединен с трофобластической оболочкой узким соединительным стеблем, который развивается в пуповину, чтобы прикрепить плаценту к эмбриону. [11] [15] Артерии в децидуальной оболочке ремоделируются, чтобы увеличить приток материнской крови в межворсинчатые пространства плаценты, обеспечивая газообмен и передачу питательных веществ эмбриону. Продукты жизнедеятельности эмбриона будут диффундировать через плаценту.
По мере того, как синцитиотрофобласт начинает проникать в стенку матки, также развивается внутренняя клеточная масса (эмбриобласт). Внутренняя клеточная масса является источником эмбриональных стволовых клеток , которые являются плюрипотентными и могут развиться в любую из трех клеток зародышевого слоя, и которые обладают потенциалом дать начало всем тканям и органам.
Эмбриобласт образует эмбриональный диск из двух слоев, верхний слой называется эпибластом , а нижний — гипобластом . Диск натянут между тем, что станет амниотической полостью , и желточным мешком. Эпибласт прилегает к трофобласту и состоит из столбчатых клеток; гипобласт находится ближе всего к полости бластоцисты и состоит из кубических клеток. Эпибласт мигрирует от трофобласта вниз, образуя амниотическую полость, выстилка которой образована амниобластами, развившимися из эпибласта. Гипобласт проталкивается вниз и образует выстилку желточного мешка (экзоцеломической полости). Некоторые клетки гипобласта мигрируют вдоль внутренней цитотрофобластической выстилки бластоцеля, секретируя по пути внеклеточный матрикс . Эти клетки гипобласта и внеклеточный матрикс называются мембраной Гейзера (или экзоцеломической мембраной), и они покрывают бластоцель, образуя желточный мешок (или экзоцеломическую полость). Клетки гипобласта мигрируют вдоль внешних краев этого ретикулума и формируют внезародышевую мезодерму; это разрушает внезародышевую ретикулум. Вскоре в ретикулуме образуются карманы, которые в конечном итоге сливаются, образуя хорионическую полость (внезародышевый целом).



Появляется первичная полоска , линейное скопление клеток, образованное мигрирующим эпибластом, и это знаменует начало гаструляции , которая происходит примерно на семнадцатый день (3-я неделя) после оплодотворения. Процесс гаструляции реорганизует двухслойный эмбрион в трехслойный эмбрион, а также придает эмбриону его специфическую ориентацию голова к хвосту и спереди назад с помощью первичной полоски, которая устанавливает двустороннюю симметрию . Первичный узел (или примитивный узел) образуется перед первичной полоской, которая является организатором нейруляции . Первичная ямка образуется как углубление в центре первичного узла, которое соединяется с хордой, которая находится непосредственно под ним. Узел возник из эпибластов дна амниотической полости, и именно этот узел вызывает образование нервной пластинки , которая служит основой для нервной системы.
Нервная пластинка сформируется напротив первичной полоски из эктодермальной ткани, которая утолщается и уплощается в нервную пластинку. Эпибласт в этой области перемещается вниз в полоску в месте первичной ямки, где происходит процесс, называемый ингрессией , который приводит к образованию мезодермы. Эта ингрессия заставляет клетки из эпибласта перемещаться в первичную полоску в ходе эпителиально-мезенхимального перехода ; эпителиальные клетки становятся мезенхимальными стволовыми клетками, мультипотентными стромальными клетками, которые могут дифференцироваться в различные типы клеток. Гипобласт отталкивается с пути и продолжает формировать амнион . Эпибласт продолжает двигаться и образует второй слой, мезодерму. Эпибласт теперь дифференцировался в три зародышевых листка эмбриона, так что двухслойный диск теперь является трехслойным диском, гаструлой .
Три зародышевых слоя — эктодерма , мезодерма и энтодерма — образуются в виде трех перекрывающихся плоских дисков. Именно из этих трех слоев все структуры и органы тела будут получены посредством процессов сомитогенеза , гистогенеза и органогенеза . [16] Эмбриональная энтодерма образуется путем инвагинации эпибластических клеток, которые мигрируют в гипобласт, в то время как мезодерма образуется клетками, которые развиваются между эпибластом и энтодермой. В целом, все зародышевые слои будут происходить из эпибласта. [11] [15] Верхний слой эктодермы даст начало внешнему слою кожи, центральной и периферической нервной системы , глаз , внутреннего уха и многих соединительных тканей . [17] Средний слой мезодермы даст начало сердцу и началу кровеносной системы , а также костям , мышцам и почкам . Внутренний слой энтодермы послужит отправной точкой для развития легких , кишечника , щитовидной железы , поджелудочной железы и мочевого пузыря .
После ингрессии, бластопор развивается там, где клетки ингрессировали, с одной стороны эмбриона, и он углубляется, чтобы стать архентероном , первой стадией формирования кишечника . Как и у всех вторичноротых , бластопор становится анусом , в то время как кишечник проходит через эмбрион на другую сторону, где отверстие становится ртом. С функционирующей пищеварительной трубкой гаструляция теперь завершена, и может начаться следующая стадия нейруляции .


После гаструляции эктодерма дает начало эпителиальной и нервной ткани , а гаструла теперь называется нейрулой . Нервная пластинка , которая образовалась как утолщенная пластинка из эктодермы, продолжает расширяться, и ее концы начинают складываться вверх как нервные складки . Нейруляция относится к этому процессу складывания, при котором нервная пластинка трансформируется в нервную трубку , и это происходит в течение четвертой недели. Они складываются вдоль неглубокой нервной бороздки , которая образовалась как разделительная срединная линия в нервной пластинке. Она углубляется по мере того, как складки продолжают набирать высоту, когда они встретятся и сомкнутся вместе в нервном гребне . Клетки, которые мигрируют через самую краниальную часть примитивной линии, образуют параксиальную мезодерму , которая даст начало сомитомерам , которые в процессе сомитогенеза дифференцируются в сомиты , которые сформируют склеротомы , синдетомы , [18] миотомы и дерматомы, чтобы сформировать хрящ и кость , сухожилия , дерму (кожу) и мышцы . Промежуточная мезодерма дает начало урогенитальному тракту и состоит из клеток, которые мигрируют из средней области примитивной линии. Другие клетки мигрируют через каудальную часть примитивной линии и образуют латеральную мезодерму, а те клетки, которые мигрируют через самую каудальную часть, вносят вклад в экстраэмбриональную мезодерму. [11] [15]
Эмбриональный диск сначала плоский и круглый, но в конечном итоге удлиняется, чтобы иметь более широкую головную часть и узкий каудальный конец. [10] В начале примитивная линия простирается в головном направлении и через 18 дней после оплодотворения возвращается каудально, пока не исчезнет. В головной части зародышевый слой показывает специфическую дифференциацию в начале четвертой недели, тогда как в каудальной части это происходит в конце четвертой недели. [11] Краниальные и каудальные нейропоры становятся все меньше, пока не закрываются полностью (к 26-му дню), образуя нервную трубку . [19]

Органогенез — это развитие органов , которое начинается в период с третьей по восьмую неделю и продолжается до рождения. Иногда полное развитие, как в легких, продолжается после рождения. Различные органы принимают участие в развитии многих систем органов тела.
Кроветворные стволовые клетки , дающие начало всем клеткам крови , развиваются из мезодермы. Развитие кроветворения происходит в скоплениях клеток крови, известных как кровяные островки , в желточном мешке . Кровяные островки развиваются вне эмбриона, на пупочном пузырьке, аллантоисе, соединительном стебле и хорионе, из мезодермальных гемангиобластов .
В центре кровяного островка гемангиобласты образуют гемопоэтические стволовые клетки, которые являются предшественниками всех типов клеток крови. На периферии кровяного островка гемангиобласты дифференцируются в ангиобласты , предшественники кровеносных сосудов. [20]

Сердце является первым функциональным органом, который развивается и начинает биться и перекачивать кровь примерно на 22 день. [21] Сердечные миобласты и кровяные островки в спланхноплевральной мезенхиме по обе стороны от нервной пластинки дают начало кардиогенной области . [11] : 165 Это подковообразная область вблизи головы эмбриона. К 19 дню, после клеточного сигнала , в этой области начинают формироваться две нити в виде трубок, поскольку внутри них развивается просвет. Эти две эндокардиальные трубки растут и к 21 дню мигрируют навстречу друг другу и сливаются, образуя одну примитивную сердечную трубку, трубчатое сердце . Это становится возможным благодаря складыванию эмбриона, которое выталкивает трубки в грудную полость . [22]
Также в то же время, когда формируются эндокардиальные трубки, начинается васкулогенез (развитие кровеносной системы). Он начинается на 18-й день с дифференциации клеток в спланхноплевральной мезодерме в ангиобласты , которые развиваются в уплощенные эндотелиальные клетки. Они объединяются, образуя небольшие пузырьки, называемые ангиокистами, которые соединяются, образуя длинные сосуды, называемые ангиобластическими тяжами. Эти тяжи развиваются в проницаемую сеть сплетений при формировании сосудистой сети. Эта сеть растет за счет дополнительного почкования и прорастания новых сосудов в процессе ангиогенеза . [22] После васкулогенеза и развития ранней сосудистой сети наступает стадия ремоделирования сосудов .
Трубчатое сердце быстро образует пять отдельных областей. От головы к хвосту это воронка , сердечная луковица , примитивный желудочек , примитивное предсердие и венозный синус . Первоначально вся венозная кровь поступает в венозный синус и продвигается от хвоста к голове в артериальный ствол . Он разделится, образуя аорту и легочную артерию ; сердечная луковица разовьется в правый (примитивный) желудочек; примитивный желудочек сформирует левый желудочек; примитивное предсердие станет передними частями левого и правого предсердий и их придатками, а венозный синус разовьется в заднюю часть правого предсердия , синоатриальный узел и коронарный синус . [21]
Сердечная петля начинает формировать сердце как один из процессов морфогенеза , и это завершается к концу четвертой недели. Запрограммированная гибель клеток ( апоптоз ) на соединяющихся поверхностях позволяет произойти слиянию. [22] В середине четвертой недели венозный синус получает кровь из трех основных вен: желточной , пупочной и общей кардинальной вены .
В течение первых двух месяцев развития начинает формироваться межпредсердная перегородка . Эта перегородка разделяет примитивное предсердие на правое и левое предсердие . Сначала она начинается как серповидный кусок ткани, который растет вниз как septum primum . Серповидная форма предотвращает полное закрытие предсердий, позволяя крови перекачиваться из правого в левое предсердие через отверстие, известное как ostium primum . Оно закрывается с дальнейшим развитием системы, но прежде чем это произойдет, в верхнем предсердии начинает формироваться второе отверстие ( ostium secundum ), что позволяет продолжать шунтирование крови. [22]
Вторая перегородка ( septum secundum ) начинает формироваться справа от septum primum. Это также оставляет небольшое отверстие, foramen ovale, которое является продолжением предыдущего отверстия ostium secundum. septum primum уменьшается до небольшого лоскута, который действует как клапан foramen ovale и остается таковым до его закрытия при рождении. Между желудочками также формируется septum inferius , который развивается в мышечную межжелудочковую перегородку . [22]
Пищеварительная система начинает развиваться с третьей недели, а к двенадцатой неделе органы занимают правильное положение.
Дыхательная система развивается из зачатка легкого , который появляется в вентральной стенке передней кишки примерно на четвертой неделе развития. Зачаток легкого образует трахею и два боковых отростка, известных как зачатки бронхов, которые увеличиваются в начале пятой недели, образуя левый и правый главные бронхи . Эти бронхи в свою очередь образуют вторичные (долевые) бронхи; три справа и два слева (отражая количество долей легкого). Третичные бронхи образуются из вторичных бронхов.
В то время как внутренняя оболочка гортани берет начало из зачатка легкого , ее хрящи и мышцы берут начало из четвертой и шестой глоточных дуг . [23]
В развивающемся эмбрионе формируются три различные почечные системы: пронефрос , мезонефрос и метанефрос . Только метанефрос развивается в постоянную почку. Все три происходят из промежуточной мезодермы .
Пронефрос развивается из промежуточной мезодермы в шейной области. Он не функционирует и дегенерирует до конца четвертой недели.
Мезонефрос происходит из промежуточной мезодермы в верхнем грудном и верхнем поясничном сегментах. Образуются выводные трубочки, которые входят в мезонефральный проток , который заканчивается в клоаке . Мезонефральный проток атрофируется у самок, но участвует в развитии репродуктивной системы у самцов.
Метанефрос появляется на пятой неделе развития. Вырост мезонефрального протока, мочеточниковый зачаток , проникает в метанефрическую ткань, образуя примитивную почечную лоханку , почечные чашечки и почечные пирамиды . Также формируется мочеточник .
Между четвертой и седьмой неделями развития уроректальная перегородка разделяет клоаку на урогенитальный синус и анальный канал . Верхняя часть урогенитального синуса образует мочевой пузырь , а нижняя часть — уретру . [23]
Поверхностный слой кожи , эпидермис , образуется из эктодермы . Более глубокий слой, дерма , образуется из мезенхимы .
Формирование эпидермиса начинается на втором месяце развития, и он приобретает свою окончательную структуру в конце четвертого месяца. Эктодерма делится, образуя плоский слой клеток на поверхности, известный как перидерма. Дальнейшее деление формирует отдельные слои эпидермиса .
Мезенхима, из которой будет формироваться дерма, образуется из трех источников:

В конце четвертой недели верхняя часть нервной трубки изгибается вентрально, образуя головной изгиб на уровне будущего среднего мозга — мезэнцефалона . [24] Над мезэнцефалоном находится прозэнцефалон (будущий передний мозг), а под ним — ромбовидный мозг (будущий задний мозг).
Клетки черепного нервного гребня мигрируют в глоточные дуги в качестве нейральных стволовых клеток , где они в процессе нейрогенеза развиваются в нейроны .
Зрительный пузырек ( который в конечном итоге становится зрительным нервом , сетчаткой и радужной оболочкой ) формируется на базальной пластинке переднего мозга. Крыловидная пластинка переднего мозга расширяется, образуя полушария головного мозга (теленцефалон), в то время как его базальная пластинка становится промежуточным мозгом. Наконец, зрительный пузырек растет, образуя оптический вырост.
С третьей по восьмую неделю развиваются лицо и шея .
Внутреннее ухо , среднее ухо и наружное ухо имеют различное эмбриональное происхождение.
Примерно на 22 день развития эктодерма с каждой стороны ромбовидного мозга утолщается, образуя слуховые плакоды . Эти плакоды инвагинируют , образуя слуховые ямки , а затем слуховые пузырьки . Затем слуховые пузырьки формируют вентральный и дорсальный компоненты.
Вентральный компонент образует мешочек и улитковый проток . На шестой неделе развития появляется улитковый проток и проникает в окружающую мезенхиму , перемещаясь по спирали, пока не образует 2,5 оборота к концу восьмой недели. Мешочек является оставшейся частью вентрального компонента. Он остается соединенным с улитковым протоком через узкий ductus reuniens .
Дорсальный компонент образует маточку и полукружные каналы .
Первый глоточный карман удлиняется и расширяется, образуя трубо-барабанное углубление . Это углубление дифференцируется, образуя большую часть барабанной полости среднего уха и всю евстахиеву или слуховую трубу . Узкая слуховая труба соединяет барабанную полость с глоткой . [25]
Кости среднего уха, слуховые косточки , происходят из хрящей глоточных дуг . Молоточек и наковальня происходят из хряща первой глоточной дуги , тогда как стремечко происходит из хряща второй глоточной дуги .
Наружный слуховой проход развивается из дорсальной части первой глоточной щели . Шесть ушных бугорков, которые представляют собой мезенхимальные разрастания на дорсальных поверхностях первой и второй глоточных дуг, образуют ушную раковину уха. [23]
Глаза начинают развиваться с третьей по десятую неделю.

В конце четвертой недели начинается развитие конечностей . Зачатки конечностей появляются на вентролатеральной стороне тела. Они состоят из внешнего слоя эктодермы и внутренней части, состоящей из мезенхимы , которая происходит из париетального слоя латеральной пластинки мезодермы . Эктодермальные клетки на дистальном конце зачатков образуют апикальный эктодермальный гребень , который создает область быстро пролиферирующих мезенхимальных клеток, известную как зона прогресса . Хрящ (некоторые из которых в конечном итоге становятся костями ) и мышцы развиваются из мезенхимы. [23]
Токсическое воздействие в эмбриональном периоде может стать причиной серьезных врожденных пороков развития , поскольку в это время развиваются зачатки основных систем органов.
Каждая клетка предимплантационного эмбриона имеет потенциал для формирования всех различных типов клеток в развивающемся эмбрионе. Эта клеточная потенция означает, что некоторые клетки могут быть удалены из предимплантационного эмбриона, а оставшиеся клетки будут компенсировать их отсутствие. Это позволило разработать метод, известный как предимплантационная генетическая диагностика , при котором небольшое количество клеток из предимплантационного эмбриона, созданного с помощью ЭКО , может быть удалено с помощью биопсии и подвергнуто генетической диагностике. Это позволяет отбирать эмбрионы, которые не затронуты определенными генетическими заболеваниями, и затем переносить их в матку матери .
Крестцово-копчиковые тератомы , опухоли, образованные из различных типов тканей, которые могут образовываться, как полагают, связаны с остатками примитивной полоски, которые обычно исчезают. [10] [11] [13]
Синдромы первой глоточной дуги — это врожденные нарушения деформаций лица, вызванные нарушением миграции клеток нервного гребня в первую глоточную дугу.
Spina bifida — врожденное заболевание , возникающее в результате неполного закрытия нервной трубки.
Вертикально передающиеся инфекции могут передаваться от матери к нерожденному ребенку на любой стадии его развития .
Гипоксия — состояние недостаточного снабжения плода кислородом — может стать серьезным последствием преждевременных или преждевременных родов.






