Змеи — это удлиненные, безногие рептилии подотряда Serpentes ( / s ɜːr ˈ p ɛ n t iː z / ). [2] Как и все другие чешуйчатые , змеи — это эктотермные , амниотные позвоночные , покрытые перекрывающейся чешуей . Многие виды змей имеют черепа с несколькими дополнительными суставами, чем их предки- ящерицы , что позволяет им глотать добычу, намного большую, чем их голова ( краниальный кинез ). Чтобы приспособить их узкие тела, парные органы змей (например, почки) появляются один перед другим, а не рядом друг с другом, и у большинства есть только одно функциональное легкое . Некоторые виды сохраняют тазовый пояс с парой рудиментарных когтей по обе стороны от клоаки . Ящерицы независимо эволюционировали с удлиненными телами без конечностей или со значительно редуцированными конечностями по крайней мере двадцать пять раз посредством конвергентной эволюции , что привело к появлению многих линий безногих ящериц . [3] Они напоминают змей, но у нескольких распространенных групп безногих ящериц есть веки и наружные уши, которых нет у змей, хотя это правило не является универсальным (см. Amphisbaenia , Dibamidae и Pygopodidae ).
Живые змеи встречаются на всех континентах, кроме Антарктиды, и на большинстве небольших участков суши; исключения включают некоторые крупные острова, такие как Ирландия, Исландия, Гренландия и острова Новой Зеландии, а также множество мелких островов Атлантического и центрального Тихого океанов. [4] Кроме того, морские змеи широко распространены по всему Индийскому и Тихому океанам. В настоящее время известно около тридцати семейств , включающих около 520 родов и около 3900 видов . [5] Они варьируются по размеру от крошечной, длиной 10,4 см (4,1 дюйма) барбадосской нитевидной змеи [6] до сетчатого питона длиной 6,95 метра (22,8 фута). [7] Ископаемый вид Titanoboa cerrejonensis имел длину 12,8 метра (42 фута). [8] Считается, что змеи произошли от роющих или водных ящериц, возможно, в юрский период, самые ранние известные окаменелости датируются 143–167 млн лет назад. [9] [10] Разнообразие современных змей появилось в эпоху палеоцена ( ок. 66–56 млн лет назад, после мел-палеогенового вымирания ). Самые старые сохранившиеся описания змей можно найти в Бруклинском папирусе .
Большинство видов змей неядовитые, а те, у которых есть яд, используют его в первую очередь для убийства и усмирения добычи, а не для самообороны. Некоторые обладают ядом, который достаточно силен, чтобы причинить болезненную травму или смерть человеку. Неядовитые змеи либо заглатывают добычу живьем, либо убивают ее путем удушения .
Английское слово snake происходит от древнеанглийского snaca , которое, в свою очередь, происходит от протогерманского * snak-an- ( ср. германское Schnake «кольцевая змея», шведское snok «травяная змея»), от протоиндоевропейского корня * (s)nēg-o- «ползать, ползать», который также дал sneak , а также санскритское nāgá «змея». [11] Слово вытеснило adder , поскольку adder продолжало сужаться в значении, хотя в древнеанглийском næddre было общим словом для змеи. [12] Другой термин, serpent , происходит от французского, в конечном счете от индоевропейского * serp- «ползать», [13] который также дал древнегреческое ἕρπω ( hérpō ) «я ползаю» и санскритское sarpá «змея». [14]
Все современные змеи сгруппированы в подотряд Serpentes в таксономии Линнея , часть отряда Squamata , хотя их точное размещение среди чешуйчатых остается спорным. [15]
Два инфраотряда Serpentes — Alethinophidia и Scolecophidia . [15] Это разделение основано на морфологических характеристиках и сходстве последовательностей митохондриальной ДНК . Alethinophidia иногда разделяют на Henophidia и Caenophidia , причем последняя состоит из «колуброидных» змей ( колубрид , гадюк , элапидов , гидрофиид и атрактаспид ) и акрохордид, в то время как другие семейства алетинофид включают Henophidia. [16] Хотя сегодня они не сохранились, Madtsoiidae , семейство гигантских, примитивных, похожих на питона змей, существовало примерно до 50 000 лет назад в Австралии, представленное такими родами, как Wonambi . [17]
Недавние молекулярные исследования подтверждают монофилию клад современных змей, сколекофидий, тифлопидий + аномалепидий, алетинофидий, основных алетинофидий, уропелтид ( Cylindrophis , Anomochilus , uropeltines), макростомат, боидов, боидов, питонид и ценофидий. [18]
В то время как змеи являются безногими рептилиями, эволюционировавшими от ящериц (и сгруппированными с ними), существует много других видов ящериц, которые потеряли свои конечности независимо, но которые внешне похожи на змей. К ним относятся верблюд-медведица , стеклянная змея и амфисбены . [22]
Ископаемая летопись змей относительно бедна, поскольку скелеты змей , как правило, небольшие и хрупкие, что делает окаменение редким. Ископаемые останки, легко идентифицируемые как змеи (хотя часто сохраняющие задние конечности), впервые появляются в ископаемой летописи в меловой период. [24] Самые ранние известные настоящие ископаемые змеи (члены коронной группы Serpentes) происходят от морских симолиофиид , старейшим из которых является позднемеловой ( сеноманский ярус) Haasiophis terrasanctus с Западного берега , [1] возраст которого составляет от 112 до 94 миллионов лет. [25]
На основании геномного анализа можно с уверенностью утверждать, что змеи произошли от ящериц . [23] Этот вывод также подтверждается сравнительной анатомией и палеонтологической летописью. [26] : 11 [27] [23]
Питоны и удавы — примитивные группы среди современных змей — имеют рудиментарные задние конечности: крошечные когтистые пальцы, известные как анальные шпоры , которые используются для хватания во время спаривания. [26] : 11 [28] Семейства Leptotyphlopidae и Typhlopidae также обладают остатками тазового пояса, которые при визуальном осмотре выглядят как роговые выступы.
Передние конечности отсутствуют у всех известных змей. Это вызвано эволюцией их Hox-генов , контролирующих морфогенез конечностей . Осевой скелет общего предка змей, как и у большинства других четвероногих, имел региональную специализацию, состоящую из шейных, грудных, поясничных, крестцовых и хвостовых позвонков. В начале эволюции змей экспрессия Hox-генов в осевом скелете, ответственная за развитие грудной клетки, стала доминирующей. В результате все позвонки, расположенные впереди зачатков задних конечностей (если они присутствуют), имеют одинаковую торакальную идентичность (за исключением атланта , оси и 1–3 шейных позвонков). Другими словами, большая часть скелета змеи представляет собой чрезвычайно расширенную грудную клетку. Ребра находятся исключительно на грудных позвонках. Шейные, поясничные и тазовые позвонки сильно редуцированы (присутствует только 2–10 поясничных и тазовых позвонков), в то время как от хвостовых позвонков остался только короткий хвост. Тем не менее, хвост все еще достаточно длинный, чтобы иметь важное значение у многих видов, и модифицирован у некоторых водных и древесных видов. [ необходима цитата ]
Многие современные группы змей возникли в палеоцене , наряду с адаптивной радиацией млекопитающих после вымирания (нептичьих) динозавров . Расширение лугов в Северной Америке также привело к взрывной радиации среди змей. [29] Ранее змеи были незначительным компонентом североамериканской фауны, но в миоцене количество видов и их распространенность резко возросли с первым появлением гадюк и аспид в Северной Америке и значительным разнообразием ужеобразных (включая происхождение многих современных родов, таких как Nerodia , Lampropeltis , Pituophis и Pantherophis ). [29]
Существуют ископаемые свидетельства, позволяющие предположить, что змеи могли произойти от роющих ящериц [30] в меловой период . [31] Ранний ископаемый родственник змей, Najash rionegrina , был двуногим роющим животным с крестцом и был полностью наземным . [32] Najash , который жил 95 миллионов лет назад, также имел череп с несколькими чертами, типичными для ящериц, но развил некоторые подвижные суставы черепа, которые определяют гибкий череп у большинства современных змей. Вид не показал никакого сходства с современными роющими слепыми змеями, которые часто рассматриваются как самая примитивная группа существующих форм. [33] Одним из существующих аналогов этих предполагаемых предков является безухий варан Lanthanotus с Борнео (хотя он также является полуводным ). [34] Подземные виды развили тела, обтекаемые для рытья нор, и в конечном итоге потеряли свои конечности. [34] Согласно этой гипотезе, такие особенности, как прозрачные , сросшиеся веки ( brille ) и потеря внешних ушей, эволюционировали, чтобы справиться с трудностями добывания ископаемых , такими как царапины на роговице и грязь в ушах. [31] [34] Известно, что некоторые примитивные змеи обладали задними конечностями, но их тазовые кости не имели прямого соединения с позвонками. К ним относятся ископаемые виды, такие как Haasiophis , Pachyrhachis и Eupodophis , которые немного старше Najash . [28]
Эта гипотеза была подкреплена в 2015 году открытием 113-миллионной окаменелости четвероногой змеи в Бразилии, которая была названа Tetrapodophis amplectus . У нее много змееподобных черт, она приспособлена для рытья нор, а ее желудок указывает на то, что она охотилась на других животных. [35] В настоящее время неизвестно, является ли Tetrapodophis змеей или другим видом из отряда чешуйчатых , поскольку змееподобное тело независимо эволюционировало по крайней мере 26 раз. У Tetrapodophis нет отличительных черт змеи в позвоночнике и черепе. [36] [37] Исследование, проведенное в 2021 году, помещает животное в группу вымерших морских ящериц мелового периода, известных как долихозавры , и не имеющих прямого отношения к змеям. [38]
Альтернативная гипотеза, основанная на морфологии , предполагает, что предки змей были связаны с мозазаврами — вымершими водными рептилиями мелового периода — образуя кладу Pythonomorpha . [27] Согласно этой гипотезе, считается, что сросшиеся прозрачные веки змей эволюционировали для борьбы с морскими условиями (потеря воды роговицей через осмос), а внешние уши были утрачены из-за неиспользования в водной среде. Это в конечном итоге привело к появлению животного, похожего на современных морских змей . В позднем меловом периоде змеи повторно колонизировали сушу и продолжили диверсифицироваться в современных змей. Ископаемые останки змей известны из ранних позднемеловых морских отложений, что согласуется с этой гипотезой; особенно потому, что они старше наземной Najash rionegrina . Схожая структура черепа, редуцированные или отсутствующие конечности и другие анатомические особенности, обнаруженные как у мозазавров, так и у змей, приводят к положительной кладистической корреляции, хотя некоторые из этих особенностей являются общими с варанидами. [ необходима ссылка ]
Генетические исследования последних лет показали, что змеи не так тесно связаны с варанами, как когда-то считалось, и, следовательно, не с мозазаврами, предполагаемым предком в водном сценарии их эволюции. Однако больше доказательств связывают мозазавров со змеями, чем с варанидами. Фрагментированные останки, найденные в юрском и раннем меловом периодах, указывают на более глубокие ископаемые летописи для этих групп, что может потенциально опровергнуть любую из гипотез. [39] [40]
Ископаемые останки, и филогенетические исследования показывают, что змеи произошли от ящериц , поэтому возник вопрос, какие генетические изменения привели к потере конечностей у предка змеи. Потеря конечностей на самом деле очень распространена среди современных рептилий и случалась десятки раз у сцинков , ангвидов и других ящериц. [41]
В 2016 году два исследования сообщили, что потеря конечностей у змей связана с мутациями ДНК в зоне регуляторной последовательности поляризующей активности (ZRS), регуляторной области гена sonic hedgehog , которая критически необходима для развития конечностей. У более продвинутых змей нет остатков конечностей, но у базальных змей, таких как питоны и удавы, есть следы сильно редуцированных, рудиментарных задних конечностей. У эмбрионов питонов даже полностью развиты зачатки задних конечностей, но их дальнейшее развитие останавливается мутациями ДНК в ZRS. [42] [43] [44] [45]
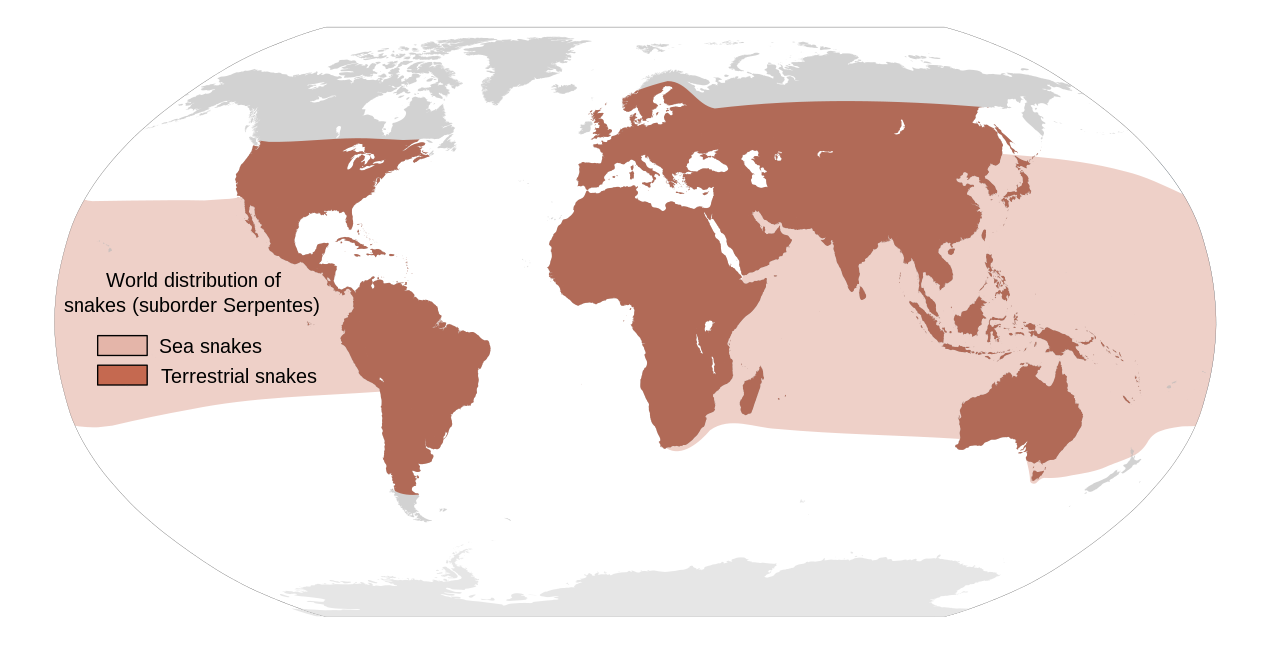
Существует около 3900 видов змей [46], которые распространены на севере вплоть до Полярного круга в Скандинавии и на юге через Австралию. [27] Змей можно найти на всех континентах, кроме Антарктиды, а также в море и на высоте до 16 000 футов (4900 м) в Гималайских горах Азии. [27] [47] : 143 Существует множество островов, на которых змеи отсутствуют, например, Ирландия , Исландия и Новая Зеландия [4] [47] (хотя северные воды Новой Зеландии нечасто посещают желтобрюхая морская змея и полосатый морской крайт ). [48]

Длина ныне вымершей Titanoboa cerrejonensis составляла 12,8 м (42 фута). [8] Для сравнения, крупнейшими из ныне живущих змей являются сетчатый питон , длина которого составляет около 6,95 м (22,8 фута), [7] и зеленая анаконда , длина которой составляет около 5,21 м (17,1 фута), и которая считается самой тяжелой змеей на Земле — ее вес составляет 97,5 кг (215 фунтов). [49]
На другом конце шкалы находится самая маленькая из ныне существующих змей — Leptotyphlops carlae , длиной около 10,4 см (4,1 дюйма). [6] Большинство змей — довольно мелкие животные, длиной около 1 м (3,3 фута). [50]
Некоторые из наиболее высокоразвитых сенсорных систем обнаружены у змей семейства Crotalidae, или ямкоголовых змей — гремучих змей и их сородичей. У ямкоголовых змей есть все органы чувств других змей, а также дополнительные вспомогательные средства. Яма относится к особым инфракрасно-чувствительным рецепторам, расположенным по обе стороны головы, между ноздрями и глазами. Фактически, ямка выглядит как дополнительная пара ноздрей. Все змеи обладают способностью ощущать тепло с помощью осязательных и тепловых рецепторов, как и другие животные; однако высокоразвитая ямка ямкоголовых змей отличается. Каждая ямка состоит из полости ямки и внутренней полости, большая из них находится сразу за и, как правило, ниже уровня ноздри и открывается вперед. За этой большей полостью находится более тонкая, едва заметная; полости соединены внутри, разделенные только мембраной с нервами, которые необычайно настроены на обнаружение изменений температуры между. Как и в перекрывающихся полях зрения человеческих глаз, обращенные вперед ямки по обе стороны лица в совокупности создают поле зрения: ямкоголовая змея может различать объекты и их окружение, а также точно оценивать расстояние между объектами и собой. Теплочувствительная способность ямкоголовой змеи настолько велика, что она может реагировать на разницу всего в одну треть градуса по Фаренгейту. Другие змеи, чувствительные к инфракрасному излучению, имеют несколько более мелких губных ямок, выстилающих верхнюю губу, чуть ниже ноздрей. [51]
Змея выслеживает свою добычу с помощью обоняния, собирая частицы в воздухе своим раздвоенным языком , а затем передавая их в вомероназальный орган или орган Якобсона во рту для исследования. [51] Развилка в языке обеспечивает своего рода направленное обоняние и вкус одновременно. [51] Язык змеи постоянно находится в движении, отбирая частицы из воздуха, земли и воды, анализируя найденные химические вещества и определяя наличие добычи или хищников в местной среде. У водных змей, таких как анаконда , язык эффективно функционирует под водой. [51]
Чтобы уловить частицы в воздухе, язык выбрасывается. Подобно руке, которая что-то берет на вес, развилка на языке одновременно обеспечивает своего рода чувство направления. У змей хорошее обоняние, но это чувство значительно усиливается в окне особого органа, органа Якобсона. Когда язык отодвигается в рот, раздвоенный кончик вдавливается в полости органа Якобсона. Оттянутые в точку, язык и орган Якобсона работают сообща для анализа вкуса и запаха. Сам орган дает змее экстрасенсорный канал. В буквальном смысле змея получает вкус окрестностей, способная скользить в комнатах информации, как будто двери открыты. [52] [ нужна страница ] [51]

Вплоть до середины 20 века считалось, что змеи не слышат. [53] [54] На самом деле у змей есть две отдельные и полностью независимые системы. Одна из этих систем, соматическая, включает передачу частот через вентральные кожные рецепторы через позвоночник. Другая система включает вибрации, которые передаются через ослабленные легкие змеи в мозг через черепной нерв. Чувствительность змеи к вибрации чрезвычайно высока. [53] [51] [54] В тихой комнате змея может слышать, как кто-то тихо говорит. [52] [ нужна страница ]
Зрение змей сильно различается у разных видов. У некоторых из них острое зрение, а другие способны отличать только свет от тьмы, но важной тенденцией является то, что зрительное восприятие змей достаточно адекватно для отслеживания движений. [55] Как правило, зрение лучше всего у змей, живущих на деревьях, и слабее всего у змей, роющих норы. У некоторых из них бинокулярное зрение , когда оба глаза способны фокусироваться на одной и той же точке, примером этого является азиатская виноградная змея . Большинство змей фокусируются, перемещая хрусталик вперед и назад по отношению к сетчатке . Дневные змеи имеют круглые зрачки, а многие ночные змеи имеют щелевидные зрачки. Большинство видов обладают тремя зрительными пигментами и, вероятно, способны видеть два основных цвета при дневном свете. Кольчатая морская змея и род Helicops , по-видимому, восстановили большую часть своего цветового зрения в качестве адаптации к морской среде, в которой они обитают. [56] [57] Был сделан вывод, что последние общие предки всех змей имели зрение, чувствительное к УФ -излучению, но большинство змей, которые зависят от своего зрения для охоты при дневном свете, развили линзы, которые действуют как солнцезащитные очки для фильтрации УФ-излучения, что, вероятно, также обостряет их зрение за счет улучшения контрастности . [ 58] [59]
Кожа змеи покрыта чешуей . Вопреки распространенному мнению о том, что змеи скользкие (из-за возможной путаницы змей с червями ), змеиная кожа имеет гладкую, сухую текстуру. Большинство змей используют специализированные брюшные чешуйки для передвижения, что позволяет им цепляться за поверхности. Чешуя на теле может быть гладкой, килеватой или зернистой. Веки змеи представляют собой прозрачные «очковые» чешуйки, также известные как brille , которые остаются постоянно закрытыми. [ необходима цитата ]
У змеи кожа была модифицирована для ее специализированной формы передвижения. Между внутренним и внешним слоями лежит дерма, которая содержит все пигменты и клетки, составляющие отличительный рисунок и цвет змеи. Эпидермис, или внешний слой, образован из вещества, называемого кератином , которое у млекопитающих является тем же основным материалом, который образует ногти, когти и волосы. Эпидермис змеи из кератина обеспечивает ей броню, необходимую для защиты внутренних органов и уменьшения трения при прохождении по камням. Части этой кератиновой брони грубее других. Менее ограниченная часть перекрывает переднюю часть чешуи под ней. Между ними лежит сложенный назад соединительный материал, также из кератина, также являющийся частью эпидермиса. Этот сложенный назад материал дает змее волнообразные движения или поедает вещи, которые больше окружности ее тела. [52] [ нужна страница ]
Сбрасывание чешуек называется экдизисом (или, в обычном употреблении, линькой или отшелушиванием ). Змеи сбрасывают весь внешний слой кожи целиком. [60] Чешуя змеи не является дискретной, а представляет собой продолжение эпидермиса — поэтому она сбрасывается не по отдельности, а как полный внешний слой во время каждой линьки, подобно носку, вывернутому наизнанку. [61]
У змей широкое разнообразие узоров окраски кожи, которые часто связаны с поведением, например, склонностью убегать от хищников. Змеи, которые подвергаются высокому риску нападения хищников, как правило, однотонные или имеют продольные полосы, что дает хищникам мало ориентиров, что позволяет змее ускользнуть, не будучи замеченной. Однотонные змеи обычно используют активные стратегии охоты, поскольку их узор позволяет им отправлять жертве мало информации о движении. Пятнистые змеи обычно используют стратегии, основанные на засаде, вероятно, потому, что это помогает им смешиваться с окружающей средой с объектами неправильной формы, такими как палки или камни. Пятнистый узор также может помочь змеям смешиваться с окружающей средой. [62]
Форма и количество чешуек на голове, спине и животе часто характерны и используются в таксономических целях. Чешуи называются в основном в соответствии с их положением на теле. У «продвинутых» ( Caenophidian ) змей широкие чешуйки на животе и ряды спинных чешуек соответствуют позвонкам , что позволяет подсчитывать их без необходимости рассечения . [ требуется цитата ]

Линька (или «экдизис») служит нескольким целям — она позволяет заменить старую, изношенную кожу и может быть синхронизирована с циклами спаривания, как и у других животных. Линька происходит периодически в течение всей жизни змеи. Перед каждой линькой змея регулирует свой рацион и ищет надежное убежище. Непосредственно перед линькой кожа становится серой, а глаза змеи — серебристыми. Внутренняя поверхность старой кожи разжижается, заставляя ее отделяться от новой кожи под ней. Через несколько дней глаза очищаются, и змея вытягивается из старой кожи, которая раскалывается. Змея трется телом о грубые поверхности, чтобы облегчить сброс старой кожи. Во многих случаях отброшенная кожа отслаивается назад по всему телу от головы до хвоста одним куском, как будто с книги снимают суперобложку, обнажая новый, более крупный и яркий слой кожи, который образовался под ней. [61] [63] Обновление кожи путем линьки предположительно увеличивает массу некоторых животных, таких как насекомые, но в случае со змеями это оспаривается. [61] [64] Сбрасывание кожи может высвобождать феромоны и оживлять цвет и узоры кожи, что повышает привлекательность партнеров. [65]
Змеи могут линять четыре-пять раз в год, в зависимости от погодных условий, наличия пищи, возраста змеи и других факторов. [52] [ нужна страница ] [63] Теоретически возможно идентифицировать змею по ее сброшенной коже, если она достаточно цела. [61] Мифологические ассоциации змей с символами исцеления и медицины , как изображено на Жезле Асклепия , являются производными от линьки. [66]
Можно попытаться определить пол змеи, когда вид не имеет отчетливо выраженного полового диморфизма , путем подсчета чешуек. Клоака зондируется и измеряется по отношению к субкаудальным чешуйкам . [67] Подсчет чешуек определяет, является ли змея самцом или самкой, так как гемипенис зондируемого самца обычно длиннее. [67] [ необходимо разъяснение ]
_skull_1_(cropped).jpg/1280px-Reticulated_python_(Python_reticulatus)_skull_1_(cropped).jpg)
Череп змеи отличается от черепа ящерицы несколькими способами. У змей более гибкие челюсти, то есть вместо соединения верхней и нижней челюсти челюсти змеи соединены костным шарниром, который называется квадратной костью . Между двумя половинами нижней челюсти у подбородка находится эластичная связка, которая обеспечивает разделение. Это позволяет змее глотать пищу большего размера в пропорциональном отношении к своему размеру и дольше обходиться без нее, поскольку змеи съедают относительно больше за одно кормление. [68] Поскольку стороны нижней челюсти могут двигаться независимо друг от друга, змея, положившая челюсть на поверхность, имеет стереослуховое восприятие , используемое для определения положения добычи. Путь челюсть-квадратная кость-стремя способен обнаруживать колебания в ангстремном масштабе, несмотря на отсутствие наружного уха и отсутствие механизма согласования импеданса , обеспечиваемого слуховыми косточками у других позвоночных. [69] [70] В черепе змеи мозг хорошо защищен. Поскольку ткани мозга могут быть повреждены через нёбо, эта защита особенно ценна. Твердый и полный нейрокраниум змей закрыт спереди. [52] [ нужна страница ] [71]

Скелет большинства змей состоит исключительно из черепа, подъязычной кости , позвоночника и ребер, хотя змеи -генофиды сохраняют остатки таза и задних конечностей. Подъязычная кость — это небольшая кость, расположенная сзади и вентрально от черепа, в области «шейки», которая служит местом прикрепления мышц языка змеи, как и у всех других четвероногих . Позвоночник состоит из 200–400 позвонков, а иногда и больше. Каждый позвонок тела имеет по два ребра, сочленяющихся с ними. Хвостовые позвонки сравнительно немногочисленны (часто менее 20% от общего числа) и лишены ребер. Позвонки имеют выступы, которые обеспечивают прочное прикрепление мышц, что позволяет передвигаться без конечностей. [ необходима цитата ]
Каудальная аутотомия (самоампутация хвоста), особенность, обнаруженная у некоторых ящериц, отсутствует у большинства змей. [72] В редких случаях, когда она существует у змей, каудальная аутотомия является межпозвоночной (то есть разделением соседних позвонков), в отличие от ящериц, которая является внутрипозвоночной, то есть разрыв происходит вдоль предопределенной плоскости перелома, присутствующей на позвонке. [73] [74]
У некоторых змей, особенно у удавов и питонов, есть остатки задних конечностей в виде пары тазовых шпор . Эти небольшие, похожие на когти выступы по обе стороны клоаки являются внешней частью рудиментарного скелета задних конечностей, который включает остатки подвздошной кости и бедренной кости. [ необходима цитата ]
Змеи — полифиодонты с постоянно заменяемыми зубами. [75]

Змеи и другие неархозавровые ( крокодилы , динозавры + птицы и их союзники) рептилии имеют трехкамерное сердце, которое контролирует кровеносную систему через левое и правое предсердие и один желудочек. [76] Внутри желудочек разделен на три взаимосвязанные полости: артериальную полость, легочную полость и венозную полость. [77] Венозная полость получает дезоксигенированную кровь из правого предсердия, а артериальная полость получает оксигенированную кровь из левого предсердия. Под венозной полостью расположена легочная полость, которая перекачивает кровь в легочный ствол. [78]
Сердце змеи заключено в мешок, называемый перикардом , расположенный в месте разветвления бронхов . Сердце способно двигаться благодаря отсутствию диафрагмы; эта регулировка защищает сердце от потенциального повреждения, когда крупная проглоченная добыча проходит через пищевод . Селезенка прикреплена к желчному пузырю и поджелудочной железе и фильтрует кровь. Тимус , расположенный в жировой ткани над сердцем, отвечает за выработку иммунных клеток в крови. Сердечно-сосудистая система змей уникальна наличием почечной портальной системы, в которой кровь из хвоста змеи проходит через почки, прежде чем вернуться в сердце. [79]
Кровеносная система змеи в основном похожа на кровеносную систему любого другого позвонка. Однако змеи не регулируют внутреннюю температуру своей крови. Называемые холоднокровными, змеи на самом деле имеют кровь, которая реагирует на изменение температуры непосредственной окружающей среды. Змеи могут регулировать температуру крови движением. Слишком долго находясь под прямыми солнечными лучами, кровь змеи нагревается сверх допустимой нормы. Оставленная на льду или снегу, змея может замерзнуть. В умеренных зонах с выраженными сезонными изменениями змеи, живущие вместе, приспособились к натиску зимы. [52] [ нужна страница ]
Рудиментарное левое легкое часто бывает маленьким или иногда даже отсутствует, так как трубчатые тела змей требуют, чтобы все их органы были длинными и тонкими. [ 79] У большинства видов функционирует только одно легкое. Это легкое содержит васкуляризированную переднюю часть и заднюю часть, которая не функционирует в газообмене. [79] Это «мешковидное легкое» используется в гидростатических целях для регулирования плавучести у некоторых водных змей, а его функция остается неизвестной у наземных видов. [79] Многие парные органы, такие как почки или репродуктивные органы , расположены в шахматном порядке внутри тела, один впереди другого. [79]
Змея с ее особым расположением органов может достичь большей эффективности. [ по сравнению с? ] Например, легкое заключает в части, ближайшей к голове и горлу, орган потребления кислорода, в то время как другая половина используется для резерва воздуха. Расположение пищевода-желудка-кишечника представляет собой прямую линию. Она заканчивается там, где открываются кишечный, мочевой и репродуктивный тракты, в камере, называемой клоакой. [52] [ нужна страница ]
У змей нет лимфатических узлов . [79]

Кобры, гадюки и близкородственные виды используют яд , чтобы обездвижить, ранить или убить свою добычу. Яд представляет собой модифицированную слюну , доставляемую через клыки . [26] [80] : 243 Клыки «продвинутых» ядовитых змей, таких как гадюки и аспидовые, полые, что позволяет вводить яд более эффективно, а клыки змей с задними клыками, таких как бумсланг, просто имеют канавку на заднем крае, чтобы направлять яд в рану. Яды змей часто специфичны для добычи, и их роль в самообороне вторична. [26] [80] : 243
Яд, как и все слюнные секреции, является предварительным перевариванием, которое инициирует расщепление пищи на растворимые соединения, способствуя правильному пищеварению. Даже укусы неядовитых змей (как и укусы любых животных) вызывают повреждение тканей. [26] [80] : 209
Некоторые птицы, млекопитающие и другие змеи (например, королевские змеи ), которые охотятся на ядовитых змей, выработали устойчивость и даже иммунитет к определенным ядам. [26] : 243 Ядовитые змеи включают три семейства змей и не составляют формальную таксономическую классификационную группу. [ необходима ссылка ]
Разговорный термин «ядовитая змея» обычно является неверным обозначением змей. Яд вдыхается или проглатывается, тогда как яд, вырабатываемый змеями, вводится в жертву через клыки. [ 81] Однако есть два исключения: Rhabdophis изолирует токсины из жаб, которых ест, а затем выделяет их из затылочных желез, чтобы отпугивать хищников; а небольшая необычная популяция подвязочных змей в американском штате Орегон сохраняет в своей печени достаточно токсинов от проглоченных тритонов , чтобы быть эффективно ядовитыми для небольших местных хищников (таких как вороны и лисы ). [82]
Яды змей представляют собой сложные смеси белков , [80] и хранятся в ядовитых железах на затылке. [82] У всех ядовитых змей эти железы открываются через протоки в бороздчатые или полые зубы в верхней челюсти. [26] : 243 [81] Белки потенциально могут быть смесью нейротоксинов (которые поражают нервную систему), гемотоксинов (которые поражают кровеносную систему), цитотоксинов (которые напрямую поражают клетки), бунгаротоксинов (родственных нейротоксинам, но также напрямую воздействующих на мышечную ткань) и многих других токсинов, которые по-разному влияют на организм. [81] [80] Почти весь змеиный яд содержит гиалуронидазу , фермент, который обеспечивает быструю диффузию яда. [26] : 243
Ядовитые змеи, использующие гемотоксины, обычно имеют клыки в передней части рта, что облегчает им введение яда в своих жертв. [80] [81] Некоторые змеи, использующие нейротоксины (например, мангровая змея ), имеют клыки в задней части рта, причем клыки загнуты назад. [83] Это затрудняет как использование змеей своего яда, так и их доение учеными. [81] Однако аспидовые змеи, такие как кобры и крайты, являются протероглифами — у них полые клыки, которые не могут быть подняты к передней части рта, и они не могут «нанести удар», как гадюка. Они должны фактически укусить жертву. [26] : 242
Было высказано предположение, что все змеи могут быть ядовитыми в определенной степени, при этом безвредные змеи имеют слабый яд и не имеют клыков. [84] Согласно этой теории, большинство змей, которые помечены как «неядовитые», будут считаться безвредными, потому что у них либо отсутствует способ доставки яда, либо они не способны доставить достаточно яда, чтобы подвергнуть опасности человека. Теория постулирует, что змеи могли произойти от общего предка ящерицы, который был ядовитым, а также что ядовитые ящерицы, такие как ядозубы , ящерицы -бусины , вараны и ныне вымершие мозазавры , могли произойти от этого же общего предка. Они разделяют эту « ядовитую кладу » с различными другими видами ящеров . [ необходима цитата ]
Ядовитые змеи подразделяются на два таксономических семейства:
Существует третье семейство, включающее опистоглифных (с задним клыком) змей (а также большинство других видов змей):
Хотя змеи используют широкий спектр репродуктивных способов, все они используют внутреннее оплодотворение . Это достигается с помощью парных раздвоенных гемипенов , которые хранятся, перевернутые, в хвосте самца. [85] Гемипены часто имеют бороздки, крючки или шипы — они предназначены для захвата стенок клоаки самки . [86] [85] Клитор самки змеи состоит из двух структур, расположенных между клоакой и пахучими железами. [87]
Большинство видов змей откладывают яйца , которые они бросают вскоре после откладывания. Однако некоторые виды (например, королевская кобра) строят гнезда и остаются поблизости от вылупившихся детенышей после инкубации. [85] Большинство питонов обвиваются вокруг своих кладок и остаются с ними до тех пор, пока они не вылупятся. [88] Самка питона не покидает яйца, за исключением случаев, когда она греется на солнце или пьет воду. Она даже «дрожит», чтобы вырабатывать тепло для инкубации яиц. [88]
Некоторые виды змей являются яйцеживородящими и сохраняют яйца внутри своих тел до тех пор, пока они почти не будут готовы вылупиться. [89] [90] Несколько видов змей, такие как удав и зеленая анаконда, являются полностью живородящими , выкармливая своих детенышей через плаценту , а также желточный мешок ; это очень необычно для рептилий и обычно встречается у серых акул или плацентарных млекопитающих . [89] [90] Задержка яиц и живорождение чаще всего связаны с более холодной средой. [85] [90]

Половой отбор у змей демонстрируется 3000 видами, каждый из которых использует разные тактики для получения партнеров. [91] Ритуальный бой между самцами за самок, с которыми они хотят спариться, включает в себя топпинг, поведение, демонстрируемое большинством гадюковых, при котором один самец обвивается вокруг вертикально поднятой передней части тела своего противника и заставляет его опуститься. Обычно кусание шеи происходит, когда змеи переплетаются. [92]
Партеногенез — это естественная форма размножения, при которой рост и развитие эмбрионов происходят без оплодотворения. Agkistrodon contortrix (медянка) и Agkistrodon piscivorus (хлопковая змея) могут размножаться факультативным партеногенезом , что означает, что они способны переключаться с полового способа размножения на бесполый способ. [93] Наиболее вероятным типом партеногенеза является аутомиксис с терминальным слиянием, процесс, при котором два терминальных продукта одного и того же мейоза сливаются, образуя диплоидную зиготу . Этот процесс приводит к гомозиготности по всему геному , экспрессии вредных рецессивных аллелей и часто к аномалиям развития. Как рожденные в неволе, так и рожденные в дикой природе медянки и хлопковые змеи, по-видимому, способны к этой форме партеногенеза. [93]
Размножение у чешуйчатых рептилий почти исключительно половое. Самцы обычно имеют пару хромосом ZZ, определяющих пол, а самки — пару ZW. Однако колумбийский радужный удав ( Epicrates maurus ) также может размножаться факультативным партеногенезом, что приводит к появлению потомства самок WW. [94] Самки WW, вероятно, производятся путем терминального аутомиксиса. [ необходима цитата ]

Эмбриональное развитие змеи изначально следует тем же этапам, что и любой эмбрион позвоночного . Эмбрион змеи начинается как зигота , подвергается быстрому делению клеток, образует зародышевый диск , также называемый бластодиском, затем подвергается гаструляции , нейруляции и органогенезу . [96] Деление клеток и пролиферация продолжаются до тех пор, пока не разовьется ранний эмбрион змеи и не будет наблюдаться типичная форма тела змеи. [96] Множественные особенности отличают эмбриологическое развитие змей от других позвоночных, двумя значимыми факторами являются удлинение тела и отсутствие развития конечностей. [ необходима ссылка ]

Удлинение тела змеи сопровождается значительным увеличением количества позвонков (у мышей 60 позвонков, тогда как у змей их может быть более 300). [95] Это увеличение позвонков происходит из-за увеличения сомитов во время эмбриогенеза, что приводит к увеличению количества развивающихся позвонков. [95] Сомиты образуются в пресомитной мезодерме из-за набора осцилляционных генов, которые направляют часы сомитогенеза . Часы сомитогенеза змеи работают с частотой в 4 раза выше, чем у мыши (после коррекции на время развития), создавая больше сомитов и, следовательно, создавая больше позвонков. [95] Считается, что эта разница в скорости часов вызвана различиями в экспрессии гена Lunatic fringe , гена, участвующего в часах сомитогенеза. [97]
Существует обширная литература, посвященная развитию/отсутствию развития конечностей у эмбрионов змей и экспрессии генов, связанной с различными стадиями. У базальных змей , таких как питон, эмбрионы на ранних стадиях развития демонстрируют зачаток задней конечности , который развивается с некоторым хрящом и хрящевым элементом таза, однако он дегенерирует перед вылуплением. [98] Это наличие рудиментарного развития предполагает, что некоторые змеи все еще подвергаются редукции задних конечностей, прежде чем они будут устранены. [99] Нет никаких доказательств у базальных змей зачатков передних конечностей и нет примеров инициации зачатков передних конечностей змей у эмбриона, поэтому мало что известно об утрате этого признака. [99] Недавние исследования показывают, что редукция задних конечностей может быть вызвана мутациями в энхансерах гена SSH , [99] однако другие исследования показали, что мутации в генах Hox или их энхансерах могут способствовать отсутствию конечностей у змей. [95] Поскольку многочисленные исследования обнаружили доказательства, указывающие на то, что различные гены играют роль в потере конечностей у змей, вполне вероятно, что множественные мутации генов имели аддитивный эффект, приводящий к потере конечностей у змей [100]


В регионах, где зимы слишком холодные для змей, чтобы выдерживать их, оставаясь активными, местные виды вступают в период спячки . В отличие от спячки , во время которой спящие млекопитающие на самом деле спят, рептилии, находящиеся в состоянии спячки, бодрствуют, но неактивны. Отдельные змеи могут зимовать в норах, под грудами камней или внутри упавших деревьев, или большое количество змей может собираться вместе в спячку . [ требуется цитата ]



Все змеи строго плотоядны , охотясь на мелких животных, включая ящериц, лягушек, других змей, мелких млекопитающих, птиц, яйца, рыбу, улиток, червей и насекомых. [26] : 81 [27] [101] Змеи не могут кусать или разрывать свою пищу на куски, поэтому должны глотать свою добычу целиком. Пищевые привычки змеи во многом зависят от размера тела; более мелкие змеи едят более мелкую добычу. Молодые питоны могут начать питаться ящерицами или мышами, а затем перейти на небольших оленей или антилоп, став взрослыми, например. [ необходима цитата ]
Челюсть змеи представляет собой сложную структуру. Вопреки распространенному мнению, что змеи могут вывихивать челюсти, у них чрезвычайно гибкая нижняя челюсть , две половины которой не жестко соединены, и множество других суставов в черепе, которые позволяют змее открывать рот достаточно широко, чтобы проглотить добычу целиком, даже если она больше в диаметре, чем сама змея. [101] Например, африканская яйцеедящая змея имеет гибкие челюсти, приспособленные для поедания яиц, намного больших, чем диаметр ее головы. [26] : 81 У этой змеи нет зубов, но есть костные выступы на внутреннем крае позвоночника , которые она использует, чтобы разбить скорлупу при поедании яиц. [26] : 81
Большинство змей питаются разнообразной добычей, но у некоторых видов есть определенная специализация. Королевские кобры и австралийские полосатые змеи поедают других змей. У видов семейства Pareidae больше зубов на правой стороне рта, чем на левой, так как они в основном охотятся на улиток, а раковины обычно закручены по часовой стрелке. [26] : 184 [102] [103]
Некоторые змеи имеют ядовитый укус, который они используют, чтобы убить свою добычу перед тем, как съесть ее. [101] [104] Другие змеи убивают свою добычу, сжимая ее , [101] а некоторые проглатывают свою добычу, когда она еще жива. [26] : 81 [101]
После еды змеи впадают в спячку, чтобы позволить процессу пищеварения произойти; [67] это интенсивная деятельность, особенно после потребления крупной добычи. У видов, которые питаются только спорадически, весь кишечник переходит в сокращенное состояние между приемами пищи, чтобы сохранить энергию. Затем пищеварительная система «повышает регуляцию» до полной мощности в течение 48 часов после потребления добычи. Будучи эктотермными («холоднокровными»), окружающая температура играет важную роль в процессе пищеварения. Идеальная температура для переваривания пищи змеями составляет 30 °C (86 °F). В пищеварении змеи задействовано огромное количество метаболической энергии, например, температура поверхности тела южноамериканской гремучей змеи ( Crotalus durissus ) увеличивается на целых 1,2 °C (2,2 °F) во время процесса пищеварения. [105] Если змею потревожить после недавнего приема пищи, она часто отрыгивает свою добычу, чтобы иметь возможность избежать предполагаемой угрозы. Если процесс пищеварения не нарушен, он протекает очень эффективно: пищеварительные ферменты змеи растворяют и поглощают все, кроме волос (или перьев) и когтей добычи, которые выводятся вместе с отходами . [ необходима ссылка ]
Надувание капюшона (расширение области шеи) является визуальным отпугивающим фактором, в основном наблюдаемым у кобр (элапид), и в первую очередь контролируется мышцами ребер. [106] Надувание капюшона может сопровождаться выплевыванием яда в сторону угрожающего объекта, [107] и созданием специального звука; шипения. Исследования на содержащихся в неволе кобрах показали, что во время надувания капюшона поднимается 13–22% длины тела. [108]
Отсутствие конечностей не мешает движению змей. Они выработали несколько различных способов передвижения, чтобы справляться с определенными средами. В отличие от походки животных с конечностями, которые образуют континуум, каждый способ передвижения змей дискретен и отличен от других; переходы между способами резкие. [109] [110]

Боковая волнистость является единственным способом передвижения в воде и наиболее распространенным способом передвижения на суше. [110] В этом способе тело змеи попеременно изгибается влево и вправо, что приводит к серии движущихся назад «волн». [109] Хотя это движение кажется быстрым, змеи редко фиксировались движущимися быстрее, чем две длины тела в секунду, часто гораздо меньше. [111] Этот способ передвижения имеет ту же чистую стоимость транспортировки (калории, сжигаемые на метр перемещения), что и бег ящериц той же массы. [112]
Наземная боковая волнистость является наиболее распространенным способом наземного передвижения для большинства видов змей. [109] В этом режиме задние движущиеся волны толкают контактные точки в окружающей среде, такие как камни, ветки, неровности почвы и т. д. [109] Каждый из этих объектов окружающей среды, в свою очередь, создает силу реакции, направленную вперед и к средней линии змеи, что приводит к толчку вперед, в то время как боковые компоненты нейтрализуются. [113] Скорость этого движения зависит от плотности точек толчка в окружающей среде, при этом средняя плотность около 8 [ необходимо разъяснение ] вдоль длины змеи является идеальной. [111] Скорость волны точно такая же, как и скорость змеи, и в результате каждая точка на теле змеи следует по пути точки впереди нее, позволяя змеям перемещаться через очень густую растительность и небольшие отверстия. [113]
При плавании волны становятся больше по мере того, как они движутся вниз по телу змеи, и волна движется назад быстрее, чем змея движется вперед. [114] Тяга создается путем толкания их тела против воды, что приводит к наблюдаемому скольжению. Несмотря на общее сходство, исследования показывают, что схема активации мышц отличается при водной и наземной боковой волнообразной деформации, что оправдывает их название отдельных режимов. [115] Все змеи могут совершать боковые волнообразные движения вперед (с движущимися назад волнами), но только морские змеи были замечены за реверсированием движения (движением назад с движущимися вперед волнами). [109]

Чаще всего используется змеями отряда ужеобразных ( колубридами , аспидами и гадюками ), когда змея должна двигаться в среде, в которой отсутствуют неровности, от которых можно отталкиваться (что делает боковую волнистость невозможной), например, скользкая илистая равнина или песчаная дюна. Боковая волнистость — это модифицированная форма боковой волнистости, при которой все сегменты тела, ориентированные в одном направлении, остаются в контакте с землей, в то время как другие сегменты приподняты, что приводит к своеобразному «перекатывающему» движению. [116] [117] Боковая волнистость движется вперед, выбрасывая петлю из себя, а затем подтягиваясь за нее. Опуская голову, змея получает рычаг, выпрямляясь и прижимаясь к земле, она выдвигается вперед под углом, который делает ее готовой к следующему прыжку. Голова и петля по сути являются двумя ногами, на которых ходит змея. Тело змеи, выглядящее примерно перпендикулярно ее направлению, может сбить с толку наблюдателя, поскольку предубеждение может привести к ассоциации движения змеи с головой, которая ведет, и телом, которое следует. Кажется, что боковая змея движется вбок, но именно туда, куда змея идет, куда она хочет идти, голова дает четкое указание. Змея оставляет за собой след, который выглядит как серия крючков, один за другим. Змеи могут двигаться назад, чтобы отступить от врага, хотя обычно они этого не делают. [52] [ нужна страница ] Этот способ передвижения преодолевает скользкую природу песка или грязи, отталкиваясь только статическими частями на теле, тем самым сводя к минимуму скольжение. [116] Статичность точек контакта можно продемонстрировать на следах боковой змеи, на которых видны отпечатки каждой чешуйки живота без какого-либо размазывания. Этот способ передвижения имеет очень низкую калорийную стоимость, менее 1 ⁄ стоимости перемещения ящерицы на то же расстояние. [112] Вопреки распространенному мнению, нет никаких доказательств того, что боковое ветряное движение связано с тем, что песок горячий. [116]
Когда точки толчка отсутствуют, но недостаточно места для использования бокового вращения из-за боковых ограничений, например, в туннелях, змеи полагаются на локомоцию гармошкой. [109] [117] В этом режиме змея упирается задней частью своего тела в стену туннеля, в то время как передняя часть змеи вытягивается и выпрямляется. [116] Затем передняя часть сгибается и образует опорную точку, а задняя часть выпрямляется и тянется вперед. Этот режим передвижения медленный и очень сложный, до семи раз превышающий стоимость бокового волнообразного движения на том же расстоянии. [112] Эта высокая стоимость обусловлена повторяющимися остановками и стартами частей тела, а также необходимостью использования активных мышечных усилий для упирания в стенки туннеля. [ необходима цитата ]

Передвижение змей в древесных местообитаниях было изучено лишь недавно. [118] Находясь на ветвях деревьев, змеи используют несколько способов передвижения в зависимости от вида и текстуры коры. [118] В целом, змеи используют модифицированную форму передвижения гармошкой на гладких ветвях, но будут совершать боковые волнообразные движения, если имеются точки соприкосновения. [118] Змеи быстрее передвигаются на небольших ветвях и при наличии точек соприкосновения, в отличие от животных с конечностями, которые лучше справляются с перемещением на больших ветвях с небольшим «беспорядком». [118]
Планирующие змеи ( Chrysopelea ) Юго-Восточной Азии взлетают с кончиков веток, расправляя ребра и совершая боковые волнообразные движения, скользя между деревьями. [116] [119] [120] Эти змеи могут совершать контролируемое планирование на сотни футов в зависимости от высоты взлета и даже могут поворачиваться в воздухе. [116] [119]
Самый медленный способ передвижения змей — прямолинейное перемещение, которое также является единственным, при котором змее не нужно сгибать свое тело вбок, хотя она может делать это при повороте. [121] В этом способе чешуйки живота поднимаются и тянутся вперед, прежде чем опуститься вниз, а тело натянуть на них. Волны движения и стазиса проходят сзади, что приводит к серии ряби на коже. [121] Ребра змеи не двигаются в этом способе передвижения, и этот метод чаще всего используют крупные питоны , удавы и гадюки, когда преследуют добычу по открытой местности, поскольку движения змеи тонкие и их труднее обнаружить добыче. [116]


Змеи обычно не охотятся на людей. Если их не напугать или не ранить, большинство змей предпочитают избегать контакта и не нападают на людей. За исключением крупных констрикторов, неядовитые змеи не представляют угрозы для людей. Укус неядовитой змеи обычно безвреден; их зубы не приспособлены для разрывания или нанесения глубоких колотых ран, а скорее для хватания и удержания. Хотя при укусе неядовитой змеи существует возможность заражения и повреждения тканей, ядовитые змеи представляют для людей гораздо большую опасность. [26] : 209 Всемирная организация здравоохранения (ВОЗ) относит укус змеи к категории «другие запущенные состояния». [124]
Задокументированные случаи смерти от укусов змей редки. Несмертельные укусы ядовитых змей могут привести к необходимости ампутации конечности или ее части. Из примерно 725 видов ядовитых змей во всем мире только 250 способны убить человека одним укусом. В Австралии в среднем происходит только один смертельный укус змеи в год. В Индии регистрируется 250 000 укусов змей за один год, и регистрируется до 50 000 первоначальных смертей. [125] ВОЗ оценивает, что порядка 100 000 человек умирают каждый год в результате укусов змей, и примерно в три раза больше ампутаций и других постоянных увечий вызываются укусами змей ежегодно. [126]
Здоровье людей подвергается серьезной угрозе из-за укусов змей, особенно в районах, где наблюдается большое разнообразие змей и ограниченный доступ к медицинской помощи, таких как регион тропических лесов Амазонки в Южной Америке. [127] Укусы змей классифицируются Всемирной организацией здравоохранения (ВОЗ) как «другие запущенные состояния». [128] Хотя зарегистрировано не так много случаев смерти от укусов змей, укусы могут вызывать серьезные осложнения и постоянные нарушения. [128] Наиболее успешным лечением укусов змей по-прежнему является противоядие, которое изготавливается из змеиного яда. [128] Однако доступ к противоядию сильно различается в зависимости от местоположения, при этом сельские районы часто испытывают трудности как со стоимостью, так и с доступностью. [129] Клинические исследования, подготовка сыворотки и извлечение яда являются одними из сложных процедур, связанных с производством противоядия. [129] Разработка альтернативных методов лечения и повышение доступности и ценовой доступности противоядия имеют важное значение для снижения глобального воздействия укусов змей на население. [130]

В некоторых частях мира, особенно в Индии, заклинание змей — это придорожное шоу, исполняемое заклинателем. В таком шоу заклинатель змей несет корзину со змеей, которую он, по-видимому, зачаровывает, играя мелодии на своем похожем на флейту музыкальном инструменте, на которые змея реагирует. [131] На самом деле змея реагирует на движение флейты, а не на звук, который она издает, поскольку у змей нет внешних ушей (хотя у них есть внутренние уши). [131]
Закон о защите дикой природы 1972 года в Индии технически запрещает заклинание змей по причине снижения жестокости к животным. Другие типы заклинателей змей используют шоу змей и мангустов , где два животных устраивают имитацию боя; однако это не очень распространено, так как животные могут быть серьезно ранены или убиты. Заклинание змей как профессия вымирает в Индии из-за конкуренции со стороны современных форм развлечений и законов об охране окружающей среды, запрещающих эту практику. Многие индийцы никогда не видели заклинания змей, и это становится сказкой прошлого. [131] [132] [133] [134]
Племя ирула из Андхра-Прадеш и Тамил Наду в Индии было охотниками-собирателями в жарких, сухих равнинных лесах и практиковало искусство ловли змей на протяжении поколений. Они обладают обширными знаниями о змеях в полевых условиях. Обычно они ловят змей с помощью простой палки. Ранее ирула поймали тысячи змей для индустрии змеиной кожи. После полного запрета индустрии змеиной кожи в Индии и защиты всех змей в соответствии с Законом о защите дикой природы Индии 1972 года они сформировали кооператив ловцов змей и переключились на ловлю змей для извлечения яда, выпуская их в дикую природу после четырех извлечений. Собранный таким образом яд используется для производства спасающего жизнь противоядия, биомедицинских исследований и для других лекарственных препаратов. [135] Известно также, что ирула едят некоторых пойманных змей и очень полезны для истребления крыс в деревнях. [ требуется цитата ]
Несмотря на существование заклинателей змей, также были профессиональные ловцы или объездчики змей . Современный отлов змей включает в себя герпетолога, использующего длинную палку с V-образным концом. Некоторые ведущие телешоу, такие как Билл Хааст , Остин Стивенс , Стив Ирвин и Джефф Корвин , предпочитают ловить их голыми руками. [ требуется цитата ]

Потребление змеиной плоти и связанных с ней товаров является отражением многих культур по всему миру, особенно в азиатских странах, таких как Китай, Тайвань, Таиланд, Индонезия, Вьетнам и Камбоджа. Из-за предполагаемых полезных для здоровья свойств и свойств афродизиака змеиное мясо часто считается деликатесом и употребляется в пищу. [136] Принято пить вино, приправленное змеиной кровью, в попытке повысить мужественность и энергию. [136] Традиционная китайская медицина считает, что змеиное вино, традиционный напиток, настоянный на целых змеях, имеет лечебные свойства. [136] Истоки змеиного вина лежат в китайской культуре. Однако использование змеиных товаров создает моральные вопросы о сохранении и благополучии животных. [137] Важно обращать внимание и регулировать устойчивый сбор змей для употребления в пищу человеком, особенно в районах, где популяции змей сокращаются в результате деградации среды обитания и чрезмерной эксплуатации. [137]
В западном мире некоторые змеи содержатся в качестве домашних животных, особенно такие миролюбивые виды, как шаровидный питон и кукурузный полоз . Чтобы удовлетворить спрос, была разработана индустрия разведения в неволе . Змеи, выращенные в неволе, считаются предпочтительнее особей, пойманных в дикой природе, и, как правило, становятся лучшими домашними животными. [138] По сравнению с более традиционными типами домашних животных, змеи могут быть очень неприхотливыми в уходе домашними животными; им требуется минимальное пространство, так как большинство распространенных видов не превышают 5 футов (1,5 м) в длину, и их можно кормить относительно редко — обычно раз в пять-четырнадцать дней. Продолжительность жизни некоторых змей составляет более 40 лет, если им обеспечить надлежащий уход. [ требуется ссылка ]


В древней Месопотамии Нирах , бог-посланник Иштарана , изображался в виде змеи на кудуррусе или пограничных камнях . [139] Изображения двух переплетенных змей распространены в шумерском искусстве и неошумерском искусстве [139] и до сих пор время от времени появляются на цилиндрических печатях и амулетах вплоть до тринадцатого века до нашей эры. [139] Рогатая гадюка ( Cerastes cerastes ) появляется в касситских и новоассирийских кудуррусе [139] и упоминается в ассирийских текстах как магическое защитное существо. [139] Драконоподобное существо с рогами, телом и шеей змеи, передними ногами льва и задними ногами птицы появляется в месопотамском искусстве с аккадского периода до эллинистического периода (323 г. до н. э.–31 г. до н. э.). [139] Это существо, известное на аккадском языке как mušḫuššu , что означает «разъяренная змея», использовалось как символ для определенных божеств, а также как общая защитная эмблема. [139] Кажется, изначально оно было спутником бога подземного мира Ниназу , [139] но позже стало спутником хурритского бога бури Тишпака , а также, позднее, сына Ниназу Нингишзиды , вавилонского национального бога Мардука , бога писцов Набу и ассирийского национального бога Ашура. [139]
В египетской истории змея занимает ведущее место, наряду с нильской коброй, украшавшей корону фараона в древние времена. Ей поклонялись как одному из богов, а также ее использовали в зловещих целях: убийство противника и ритуальное самоубийство ( Клеопатра ). [140] Уроборос был известным древнеегипетским символом змеи, глотающей свой собственный хвост. [141] Предшественником уробороса был «Многоликий», [141] змей с пятью головами, который, согласно Амдуату , старейшей сохранившейся Книге загробной жизни , как говорили, обвивался вокруг тела бога солнца Ра в защитных целях. [ 141] Самое раннее сохранившееся изображение «истинного» уробороса происходит из позолоченных святилищ в гробнице Тутанхамона . [141] В первые века нашей эры уроборос был принят в качестве символа гностическими христианами [141] и глава 136 Pistis Sophia , раннего гностического текста, описывает «великого дракона, хвост которого находится во рту». [141] В средневековой алхимии уроборос стал типичным западным драконом с крыльями, ногами и хвостом. [141]
В Библии царь Аммона Нахаш , чье имя означает «Змея», изображен крайне негативно, как особенно жестокий и презренный враг древних евреев. [ необходима цитата ]

Древние греки использовали Горгонейон , изображение отвратительного лица со змеями вместо волос, как апотропеический символ для отражения зла. [142] В греческом мифе, описанном Псевдо-Аполлодором в его «Библиотеке» , Медуза была Горгоной со змеями вместо волос, чей взгляд обращал всех, кто смотрел на нее, в камень, и была убита героем Персеем . [143] [144] [145] В «Метаморфозах » римского поэта Овидия говорится , что Медуза когда-то была прекрасной жрицей Афины , которую Афина превратила в чудовище со змееволосым после того, как ее изнасиловал бог Посейдон в храме Афины. [146] В другом мифе, на который ссылается беотийский поэт Гесиод и который подробно описан Псевдо-Аполлодором, говорится, что герой Геракл убил Лернейскую Гидру , [147] [148] многоголовую змею, обитавшую в болотах Лерны . [147] [148]
В легендарном рассказе об основании Фив упоминается чудовищная змея, охраняющая источник, из которого новое поселение должно было черпать воду. В борьбе со змеей и ее убийстве все соратники основателя Кадма погибли, что привело к появлению термина « победа Кадмея » (т. е. победа, влекущая за собой собственную гибель). [149]
Три медицинских символа, связанных со змеями, которые используются и сегодня, — это Чаша Гигиеи , символизирующая фармацию, а также Кадуцей и Жезл Асклепия , которые являются символами, обозначающими медицину в целом. [66]
Одна из предложенных этимологий распространенного женского имени Линда заключается в том, что оно может происходить от древнегерманского Lindi или Linda , что означает «змея». [ требуется цитата ]
Индию часто называют страной змей, и она пропитана традициями, касающимися змей. [150] Змеям поклоняются как богам даже сегодня, и многие женщины льют молоко в змеиные ямы (несмотря на отвращение змей к молоку). [150] Кобра изображена на шее Шивы , а Вишну часто изображается спящим на семиглавой змее или в кольцах змеи. [151] В Индии также есть несколько храмов, посвященных исключительно кобрам, иногда называемых Наградж (Король змей), и считается, что змеи являются символами плодородия. Каждый год проводится индуистский фестиваль под названием Наг Панчами , в этот день почитают змей и молятся им. См. также Нага . [152]
Змея — одно из 12 небесных животных китайского зодиака , в китайском календаре . [153]
Многие древние перуанские культуры поклонялись природе. [154] Они придавали особое значение животным и часто изображали змей в своем искусстве. [155]

Змеи используются в индуизме как часть ритуального поклонения. [157] На ежегодном фестивале Наг Панчами участники поклоняются либо живым кобрам, либо изображениям Нагов . Господь Шива изображен на большинстве изображений со змеей, обвивающейся вокруг его шеи. [158] Пураническая литература включает в себя различные истории, связанные со змеями, например, говорится, что Шеша держит все планеты Вселенной на своих капюшонах и постоянно воспевает славу Вишну всеми своими ртами. Другие известные змеи в индуизме - Васуки , Такшака , Каркотака и Пингала . Термин Нага используется для обозначения сущностей, которые принимают форму больших змей в индуизме и буддизме . [159]
Змеи широко почитались во многих культурах, например, в Древней Греции , где змея считалась целителем. [160] Асклепий носил змеиную рану вокруг своего жезла, символ, который сегодня можно увидеть на многих машинах скорой помощи. [161] В иудаизме медная змея также является символом исцеления, спасения жизни человека от неминуемой смерти. [162]
В религиозных терминах змея и ягуар были, возможно, самыми важными животными в древней Мезоамерике . [163] «В состоянии экстаза лорды танцуют танец змеи; огромные спускающиеся змеи украшают и поддерживают здания от Чичен-Ицы до Теночтитлана , а науатльское слово coatl, означающее змею или близнеца, является частью основных божеств, таких как Мишкоатль , Кецалькоатль и Коатликуэ ». [164] В календарях майя и ацтеков пятый день недели был известен как День змеи. [ требуется цитата ]
В некоторых частях христианства искупительная работа Иисуса Христа сравнивается со спасением жизни человека посредством созерцания Нехуштана (медного змея). [165] Укротители змей используют змей как неотъемлемую часть церковного богослужения, чтобы продемонстрировать свою веру в божественную защиту. Однако чаще в христианстве змея изображают как представителя зла и хитрых заговоров, как это видно из описания в Книге Бытия змеи, искушающей Еву в Эдемском саду . [166] Предполагается, что Святой Патрик изгнал всех змей из Ирландии, обращая страну в христианство в V веке, что объясняет отсутствие там змей. [167]
В христианстве и иудаизме змея появляется в первой книге Библии, когда змея появляется перед Адамом и Евой и искушает их запретным плодом с Древа Познания . [166] Змея возвращается в Книге Исхода, когда Моисей превращает свой посох в змею как знак силы Бога, и позже, когда он создает Нехуштан, бронзовую змею на шесте, которая, если на нее посмотреть, исцеляла людей от укусов змей, досаждавших им в пустыне. Змея появляется в последний раз, символизируя Сатану , в Книге Откровения : «И он схватил дракона, древнего змея, который есть дьявол и сатана, и сковал его на тысячу лет». [168]
В неоязычестве и викке змея рассматривается как символ мудрости и знания. [169] Кроме того, змеи иногда ассоциируются с Гекатой , греческой богиней колдовства . [170]
Несколько соединений из змеиных ядов исследуются в качестве потенциальных средств лечения или профилактики боли, рака, артрита, инсульта, болезней сердца, гемофилии и гипертонии, а также для контроля кровотечения (например, во время операции). [171] [172] [173]




{kind=link}
{kind=link}