_with_its_prey.jpg/1280px-Polar_bear_(Ursus_maritimus)_with_its_prey.jpg)

Хищничество — это биологическое взаимодействие , при котором один организм, хищник , убивает и съедает другой организм, свою добычу . Это одно из семейства общих поведений кормления , которое включает паразитизм и микрохищничество (которые обычно не убивают хозяина ) и паразитоидизм (который всегда это делает, в конечном итоге). Оно отличается от поедания мертвой добычи, хотя многие хищники также питаются падалью ; оно пересекается с травоядностью , поскольку хищники семян и разрушительные плодоядные являются хищниками.
Хищники могут активно искать или преследовать добычу или ждать ее, часто затаившись. Когда добыча обнаружена, хищник оценивает, стоит ли нападать на нее. Это может включать засаду или преследование , иногда после выслеживания добычи. Если атака успешна, хищник убивает добычу, удаляет все несъедобные части, такие как панцирь или шипы, и съедает ее.
Хищники приспособлены и часто узкоспециализированы для охоты, с острыми чувствами, такими как зрение , слух или обоняние . Многие хищные животные , как позвоночные , так и беспозвоночные , имеют острые когти или челюсти, чтобы хватать, убивать и разделывать свою добычу. Другие адаптации включают скрытность и агрессивную мимикрию , которые повышают эффективность охоты.
Хищничество оказывает мощное избирательное воздействие на добычу, и добыча развивает антихищные адаптации, такие как предупреждающая окраска , тревожные крики и другие сигналы , камуфляж , подражание хорошо защищенным видам, а также защитные шипы и химические вещества. Иногда хищник и добыча оказываются в эволюционной гонке вооружений , цикле адаптаций и контрадаптаций. Хищничество было основным двигателем эволюции, по крайней мере, с кембрийского периода.

На самом базовом уровне хищники убивают и едят другие организмы. Однако концепция хищничества широка, определяется по-разному в разных контекстах и включает в себя широкий спектр методов питания; более того, некоторые отношения, которые приводят к смерти жертвы, не обязательно называются хищничеством. Паразитоид , такой как оса-ихневмон , откладывает яйца в или на своего хозяина; из яиц вылупляются личинки, которые поедают хозяина, и он неизбежно умирает. Зоологи обычно называют это формой паразитизма , хотя традиционно считается, что паразиты не убивают своих хозяев. Хищника можно определить как отличающегося от паразитоида тем, что у него много жертв, пойманных в течение его жизни, тогда как у личинки паразитоида есть только одна, или, по крайней мере, ее запас пищи предоставляется ей только один раз. [1] [2]

Существуют и другие сложные и пограничные случаи. Микрохищники — это мелкие животные, которые, как и хищники, питаются исключительно другими организмами; к ним относятся блохи и комары , которые потребляют кровь живых животных, и тли , которые потребляют сок живых растений. Однако, поскольку они обычно не убивают своих хозяев, их теперь часто считают паразитами. [3] [4] Животные, которые питаются фитопланктоном или скоплениями микробов , являются хищниками, поскольку они потребляют и убивают свои пищевые организмы, в то время как травоядные, которые объедают листья, таковыми не являются, поскольку их пищевые растения обычно переживают нападение. [5] Когда животные едят семена ( хищничество семенами или зерноядность ) или яйца ( хищничество яйцами ), они потребляют целые живые организмы, что по определению делает их хищниками. [6] [7] [8]
Падальщики , организмы, которые едят только уже мертвые организмы, не являются хищниками, но многие хищники, такие как шакалы и гиены, питаются падалью, когда появляется такая возможность. [9] [10] [5] Среди беспозвоночных общественные осы , такие как осы-осы, являются как охотниками, так и падальщиками других насекомых. [11]
Хотя примеры хищников среди млекопитающих и птиц хорошо известны, [12] хищники могут быть обнаружены в широком диапазоне таксонов, включая членистоногих. Они распространены среди насекомых, включая богомолов, стрекоз , златоглазок и скорпионов . У некоторых видов, таких как ольхоловка , только личинки являются хищными (взрослые особи не едят). Хищными являются пауки, а также другие наземные беспозвоночные, такие как скорпионы ; многоножки ; некоторые клещи , улитки и слизни ; нематоды ; и планарии . [13] В морской среде большинство книдарий (например, медузы , гидроиды ), гребневики (гребневики), иглокожие (например, морские звезды , морские ежи , плоские доллары и морские огурцы ) и плоские черви являются хищными. [14] Среди ракообразных хищниками являются омары , крабы , креветки и морские желуди , [15] а на ракообразных, в свою очередь , охотятся почти все головоногие моллюски (включая осьминогов , кальмаров и каракатиц ). [ 16 ]

Хищничество за семенами ограничивается млекопитающими, птицами и насекомыми, но встречается почти во всех наземных экосистемах. [8] [6] Хищничество за яйцами включает как специализированных хищников, таких как некоторые ужеобразные змеи , так и универсалов, таких как лисы и барсуки, которые случайно забирают яйца, когда находят их. [17] [18] [19]
Некоторые растения, такие как кувшинка , венерина мухоловка и росянка , являются плотоядными и поедают насекомых . [12] Методы хищничества растений сильно различаются, но часто включают в себя пищевую ловушку, механическую стимуляцию и электрические импульсы, чтобы в конечном итоге поймать и съесть свою добычу. [20] Некоторые плотоядные грибы ловят нематод, используя либо активные ловушки в виде сжимающих колец, либо пассивные ловушки с клейкими структурами. [21]
Многие виды простейших ( эукариот ) и бактерий ( прокариот ) охотятся на другие микроорганизмы; способ питания, очевидно, является древним и многократно эволюционировал в обеих группах. [22] [12] [23] Среди пресноводного и морского зоопланктона , будь то одноклеточный или многоклеточный, хищное выедание фитопланктона и более мелкого зоопланктона является обычным явлением и обнаружено у многих видов нанофлагеллят , динофлагеллят , инфузорий , коловраток , разнообразного спектра личинок животных меропланктона и двух групп ракообразных, а именно веслоногих и ветвистоусых . [24]

Чтобы питаться, хищник должен искать, преследовать и убивать свою жертву. Эти действия образуют цикл добычи пищи . [26] [27] Хищник должен решить, где искать добычу, основываясь на ее географическом распределении; и как только он обнаружил добычу, он должен оценить, преследовать ее или ждать лучшего выбора. Если он выбирает преследование, его физические возможности определяют способ преследования (например, засада или погоня). [28] [29] Поймав добычу, ему также может потребоваться затратить энергию на ее обработку (например, убить ее, удалить панцирь или шипы и проглотить ее). [25] [26]
Хищники могут выбирать режимы поиска, начиная от «сидеть и ждать» до активного или широкого поиска пищи . [30] [25] [31] [32] Метод «сидеть и ждать» наиболее подходит, если добыча плотная и подвижная, а у хищника низкие энергетические потребности. [30] Широкий поиск пищи требует больше энергии и используется, когда добыча малоподвижна или распределена редко. [28] [30] Существует континуум режимов поиска с интервалами между периодами движения от нескольких секунд до нескольких месяцев. Акулы, солнечные рыбы , насекомоядные птицы и землеройки почти всегда находятся в движении, в то время как пауки, плетущие паутину, водные беспозвоночные, богомолы и пустельги перемещаются редко. Между тем, ржанки и другие кулики , пресноводные рыбы, включая краппи , и личинки жуков-кокцинеллид (божьи коровки) попеременно активно ищут и сканируют окружающую среду. [30]

Распределение добычи часто сгруппировано, и хищники реагируют, выискивая участки , где добыча плотна, а затем ищут внутри участков. [25] Там, где еда находится в участках, таких как редкие косяки рыб в почти пустом океане, стадия поиска требует от хищника путешествовать в течение значительного времени и тратить значительное количество энергии, чтобы найти каждый участок пищи. [33] Например, чернобровый альбатрос регулярно совершает полеты за пищей на расстояние около 700 километров (430 миль), вплоть до максимального диапазона добычи пищи в 3000 километров (1860 миль) для размножающихся птиц, собирающих пищу для своего потомства. [a] [34] При наличии статической добычи некоторые хищники могут узнавать подходящие местоположения участков и возвращаться к ним через определенные промежутки времени, чтобы поесть. [33] Оптимальная стратегия добычи пищи для поиска была смоделирована с использованием теоремы о предельной ценности . [35]
Модели поиска часто кажутся случайными. Одним из таких является походка Леви , которая, как правило, включает кластеры коротких шагов с редкими длинными шагами. Она хорошо подходит для поведения самых разных организмов, включая бактерии, медоносных пчел, акул и людей-охотников-собирателей. [36] [37]

Найдя добычу, хищник должен решить, преследовать ее или продолжать поиски. Решение зависит от затрат и выгод. Птица, охотящаяся за насекомыми, тратит много времени на поиски, но поймать и съесть их можно быстро и легко, поэтому эффективная стратегия для птицы — съесть каждое вкусное насекомое, которое она найдет. Напротив, хищник, такой как лев или сокол, легко находит свою добычу, но ее поимка требует больших усилий. В этом случае хищник более избирателен. [28]
Одним из факторов, которые следует учитывать, является размер. Слишком маленькая добыча может не стоить усилий из-за количества энергии, которую она дает. Слишком большая, и ее может быть слишком трудно поймать. Например, богомол захватывает добычу передними ногами, и они оптимизированы для захвата добычи определенного размера. Богомолы неохотно нападают на добычу, которая далека от этого размера. Существует положительная корреляция между размером хищника и его добычи. [28]
Хищник может оценить участок и решить, стоит ли тратить время на поиски добычи. [25] Это может включать в себя некоторое знание предпочтений добычи; например, божьи коровки могут выбрать участок растительности, подходящий для их добычи тли . [38]
Чтобы захватить добычу, хищники используют целый спектр методов преследования, которые варьируются от открытого преследования ( хищничество преследования ) до внезапного удара по ближайшей добыче ( хищничество из засады ). [25] [39] [12] Другая стратегия между засадой и преследованием — это баллистический перехват , когда хищник наблюдает и предсказывает движение добычи, а затем соответствующим образом начинает атаку. [40]
Хищники, нападающие из засады или сидящие и выжидающие, являются плотоядными животными, которые захватывают добычу скрытно или неожиданно. У животных хищничество из засады характеризуется тем, что хищник сканирует окружающую среду из скрытого положения, пока не заметит добычу, а затем быстро выполняет фиксированную внезапную атаку. [41] [40] К позвоночным хищникам, нападающим из засады, относятся лягушки, рыбы, такие как акула-ангел , северная щука и восточная рыба-лягушка . [40] [42] [43] [44] Среди многих беспозвоночных хищников, нападающих из засады, есть пауки-ловушки и австралийские пауки-крабы на суше, а также раки-богомолы в море. [41] [45] [46] Хищники, нападающие из засады, часто строят норы, в которых могут спрятаться, улучшая маскировку за счет уменьшения поля зрения. Некоторые хищники, нападающие из засады, также используют приманки, чтобы привлечь добычу в зону поражения. [40] Движение захвата должно быть быстрым, чтобы поймать добычу, учитывая, что атака не может быть изменена после ее начала. [40]

Баллистический перехват — это стратегия, при которой хищник наблюдает за движением добычи, предсказывает ее движение, разрабатывает путь перехвата, а затем атакует добычу на этом пути. Это отличается от засадного нападения тем, что хищник корректирует свою атаку в соответствии с тем, как движется добыча. [40] Баллистический перехват включает в себя короткий период планирования, давая добыче возможность убежать. Некоторые лягушки ждут, пока змеи начнут свою атаку, прежде чем прыгать, сокращая время, доступное змее для перекалибровки своей атаки, и максимизируя угловую корректировку, которую змее необходимо будет сделать, чтобы перехватить лягушку в реальном времени. [40] Баллистические хищники включают в себя насекомых, таких как стрекозы, и позвоночных, таких как брызгуны (атакующие струей воды), хамелеоны (атакующие языком) и некоторые ужеобразные змеи . [40]
При преследовании хищники преследуют убегающую добычу. Если добыча убегает по прямой, захват зависит только от того, быстрее ли хищник, чем добыча. [40] Если добыча маневрирует, поворачиваясь во время бегства, хищник должен реагировать в реальном времени, чтобы рассчитать и следовать новому пути перехвата, например, с помощью параллельной навигации , когда он приближается к добыче. [40] Многие преследующие хищники используют камуфляж, чтобы приблизиться к добыче как можно ближе незамеченными ( выслеживание ), прежде чем начать преследование. [40] К преследующим хищникам относятся наземные млекопитающие, такие как люди, африканские дикие собаки, пятнистые гиены и волки; морские хищники, такие как дельфины, косатки и многие хищные рыбы, такие как тунец; [47] [48] хищные птицы (хищники), такие как соколы; и насекомые, такие как стрекозы . [49]
Экстремальная форма преследования — это охота на выносливость или настойчивость , при которой хищник изматывает добычу, преследуя ее на большом расстоянии, иногда часами. Этот метод используется охотниками -собирателями и псовыми, такими как африканские дикие собаки и домашние гончие. Африканская дикая собака — хищник с чрезвычайной настойчивостью, изматывающий отдельную добычу, преследуя ее на протяжении многих миль на относительно низкой скорости. [50]
Специализированной формой преследования добычи является выпадное питание усатых китов . Эти очень крупные морские хищники питаются планктоном , особенно крилем , ныряя и активно плавая в скоплениях планктона, а затем делая огромный глоток воды и фильтруя ее через свои перистые пластины усатых китов . [51] [52]
Преследующие хищники могут быть социальными , как лев и волк, которые охотятся группами, или одиночными. [2]
Как только хищник схватил добычу, он должен с ней обращаться: очень осторожно, если добыча опасна для употребления в пищу, например, если у нее есть острые или ядовитые шипы, как у многих хищных рыб. У некоторых сомов, таких как Ictaluridae, есть шипы на спине (спинной) и животе (грудной), которые фиксируются в вертикальном положении; когда сом мечется, когда его ловят, они могут пронзить рот хищника, возможно, смертельно. Некоторые питающиеся рыбой птицы, такие как скопа, избегают опасности шипов, разрывая свою добычу перед тем, как съесть ее. [53]
При социальном хищничестве группа хищников объединяется, чтобы убить добычу. Это позволяет убивать существ, более крупных, чем те, которых они могли бы одолеть в одиночку; например, гиены и волки объединяются, чтобы поймать и убить травоядных, таких больших, как буйвол, а львы даже охотятся на слонов. [54] [55] [56] Это также может сделать добычу более доступной с помощью таких стратегий, как вспугивание добычи и загон ее в меньшую область. Например, когда смешанные стаи птиц ищут корм, птицы впереди выпугивают насекомых, которых ловят птицы сзади. Длиннорылые дельфины образуют круг вокруг косяка рыб и движутся внутрь, концентрируя рыбу в 200 раз. [57] Охотясь сообща, шимпанзе могут ловить обезьян колобусов , которые легко ускользнули бы от индивидуального охотника, в то время как сотрудничающие ястребы Харриса могут ловить кроликов. [54] [58]

Хищники разных видов иногда объединяются, чтобы поймать добычу. В коралловых рифах , когда рыбы, такие как групер и коралловая форель, замечают добычу, которая им недоступна, они подают сигнал гигантским муренам , наполеоновым губанам или осьминогам . Эти хищники способны проникать в небольшие расщелины и выманивать добычу. [59] [60] Известно, что косатки помогают китобоям охотиться на усатых китов . [61]
Социальная охота позволяет хищникам захватывать более широкий спектр добычи, но с риском конкуренции за захваченную пищу. Одиночные хищники имеют больше шансов съесть то, что они поймают, ценой повышенных затрат энергии на поимку и повышенного риска того, что добыча сбежит. [62] [63] Хищники, нападающие из засады, часто охотятся в одиночку, чтобы снизить риск стать добычей самим. [64] Из 245 наземных представителей отряда Carnivora (группа, которая включает кошек, собак и медведей), 177 являются одиночными; и 35 из 37 диких кошек являются одиночными, [65] включая пуму и гепарда. [62] [2] Однако одиночная пума позволяет другим пумам разделить добычу, [66] а койот может быть как одиночным, так и социальным. [67] Другие одиночные хищники включают северную щуку, [68] пауков-волков и все тысячи видов одиночных ос среди членистоногих, [69] [70] а также многие микроорганизмы и зоопланктон . [22] [71]
Под давлением естественного отбора хищники выработали множество физических приспособлений для обнаружения, ловли, убийства и переваривания добычи. К ним относятся скорость, ловкость, скрытность, острые чувства, когти, зубы, фильтры и подходящие пищеварительные системы. [72]
Для обнаружения добычи хищники имеют хорошо развитое зрение , обоняние или слух . [12] У таких разнообразных хищников, как совы и пауки-скакуны, глаза направлены вперед, обеспечивая точное бинокулярное зрение в относительно узком поле зрения, тогда как у животных-жертв часто менее острое круговое зрение. Такие животные, как лисы, могут учуять свою добычу, даже если она скрыта под 2 футами (60 см) снега или земли. Многие хищники обладают острым слухом, а некоторые, такие как летучие мыши с эхолокацией, охотятся исключительно с помощью активного или пассивного использования звука. [73]
Хищники, включая больших кошек , хищных птиц и муравьев, имеют мощные челюсти, острые зубы или когти, которые они используют, чтобы схватить и убить свою добычу. Некоторые хищники, такие как змеи и питающиеся рыбой птицы, такие как цапли и бакланы, проглатывают свою добычу целиком; некоторые змеи могут расцеплять свои челюсти, чтобы проглотить большую добычу, в то время как питающиеся рыбой птицы имеют длинные копьевидные клювы, которые они используют, чтобы пронзать и хватать быстро движущуюся и скользкую добычу. [73] Рыбы и другие хищники развили способность раздавливать или открывать бронированные раковины моллюсков. [74]
Многие хищники обладают мощным телосложением и могут ловить и убивать животных, которые крупнее их самих; это относится как к мелким хищникам, таким как муравьи и землеройки, так и к крупным и заметно мускулистым плотоядным животным, таким как пума и лев . [73] [2] [75]
.JPG/1280px-Ursus_arctos_01_MWNH_145_(cropped).JPG)

Хищники часто узкоспециализированы в своем рационе и охотничьем поведении; например, евразийская рысь охотится только на мелких копытных . [76] Другие, такие как леопарды, являются более оппортунистическими универсалами, охотящимися по крайней мере на 100 видов. [77] [78] Специалисты могут быть высоко адаптированы к поимке своей предпочтительной добычи, тогда как универсалы могут быть лучше приспособлены к переключению на другую добычу, когда предпочтительная цель редка. Когда добыча имеет сгруппированное (неравномерное) распределение, оптимальная стратегия для хищника, как ожидается, будет более специализированной, поскольку добыча более заметна и может быть найдена быстрее; [79] это, по-видимому, верно для хищников неподвижной добычи, но сомнительно для подвижной добычи. [80]
При хищничестве, избирательном по размеру, хищники выбирают добычу определенного размера. [81] Крупная добыча может оказаться неприятной для хищника, в то время как мелкую добычу может быть трудно найти, и в любом случае она обеспечивает меньшее вознаграждение. Это привело к корреляции между размером хищников и их добычей. Размер также может служить убежищем для крупной добычи. Например, взрослые слоны относительно защищены от хищничества львов, но молодые особи уязвимы. [82]
Представители семейства кошачьих , такие как снежный барс (безлесные высокогорья), тигр (травянистые равнины, тростниковые болота), оцелот (леса), кошка-рыболов (заросли у воды) и лев (открытые равнины), маскируются с помощью окраски и ярких узоров, соответствующих их среде обитания. [83]
В агрессивной мимикрии некоторые хищники, включая насекомых и рыб, используют окраску и поведение для привлечения добычи. Самки светлячков Photuris , например, копируют световые сигналы других видов, тем самым привлекая самцов светлячков, которых они ловят и едят. [84] Цветочные богомолы — хищники-засадчики; замаскированные под цветы, такие как орхидеи , они привлекают добычу и хватают ее, когда она находится достаточно близко. [85] Рыбы-лягушки чрезвычайно хорошо замаскированы и активно заманивают свою добычу, чтобы приблизиться, используя эску , приманку на конце стержневидного придатка на голове, которым они мягко машут, имитируя небольшое животное, заглатывая добычу чрезвычайно быстрым движением, когда она оказывается в пределах досягаемости. [86]
Многие мелкие хищники, такие как кубомедуза, используют яд , чтобы подавлять свою добычу, [87] и яд также может помочь в пищеварении (как в случае с гремучими змеями и некоторыми пауками ). [88] [89] Мраморная морская змея , которая приспособилась к хищничеству за счет яиц, имеет атрофированные ядовитые железы, а ген ее трехпалого токсина содержит мутацию (делецию двух нуклеотидов ), которая его инактивирует. Эти изменения объясняются тем, что ее добычу не нужно подавлять. [90]

Несколько групп хищных рыб обладают способностью обнаруживать, отслеживать, а иногда, как у электрического ската , выводить из строя свою добычу, ощущая и генерируя электрические поля . [91] [92] [93] Электрический орган происходит из модифицированной нервной или мышечной ткани. [94]
Физиологическая адаптация к хищничеству включает способность хищных бактерий переваривать сложный полимер пептидогликана из клеточных стенок бактерий, на которых они охотятся. [23] Плотоядные позвоночные всех пяти основных классов (рыбы, земноводные, рептилии, птицы и млекопитающие) имеют более низкие относительные скорости транспорта сахара в аминокислоты, чем травоядные или всеядные, предположительно потому, что они получают много аминокислот из животных белков в своем рационе. [95]
Чтобы противостоять хищникам, добыча выработала защиту для использования на каждом этапе атаки. [96] [12] Они могут попытаться избежать обнаружения, [97] например, используя камуфляж и мимикрию . [98] Они могут обнаруживать хищников [99] и предупреждать других об их присутствии. [100] [101] Если их обнаружат, они могут попытаться избежать нападения, например, сигнализируя , что они токсичны или несъедобны , [102] [103] [104] сигнализируя, что преследование будет невыгодным, [105] [106] или образуя группы. [107] [108] Если они становятся целью, они могут попытаться отразить атаку с помощью защиты, такой как броня, иглы , несъедобность или толпа; [109] [110] [111] и они часто могут избежать нападения во время него , напугав хищника, [112] [113] [114] притворившись мертвыми , сбросив части тела , такие как хвосты, или просто убежав. [115] [116]

Хищники и добыча являются естественными врагами, и многие из их адаптаций, по-видимому, предназначены для противостояния друг другу. Например, летучие мыши обладают сложными системами эхолокации для обнаружения насекомых и другой добычи, а насекомые выработали множество защит, включая способность слышать эхолокационные сигналы. [117] [118] Многие преследующие хищники, которые бегают по земле, такие как волки, развили длинные конечности в ответ на возросшую скорость своей добычи. [119] Их адаптации были охарактеризованы как эволюционная гонка вооружений , пример коэволюции двух видов. [120] В геноцентрическом взгляде на эволюцию гены хищника и добычи можно рассматривать как конкурирующие за тело добычи. [120] Однако принцип «жизнь-обед» Докинза и Кребса предсказывает, что эта гонка вооружений асимметрична: если хищник не может поймать свою добычу, он теряет свой обед, в то время как если ему это удается, добыча теряет свою жизнь. [120]

Метафора гонки вооружений подразумевает постоянно растущие достижения в атаке и обороне. Однако эти адаптации имеют свою цену; например, более длинные ноги имеют повышенный риск перелома, [121] в то время как специализированный язык хамелеона, с его способностью действовать как снаряд, бесполезен для лакания воды, поэтому хамелеон должен пить росу с растительности. [122]
Принцип «жизнь-ужин» подвергался критике по нескольким причинам. Степень асимметрии в естественном отборе частично зависит от наследуемости адаптивных признаков. [122] Кроме того, если хищник теряет достаточно обедов, он также теряет свою жизнь. [121] [122] С другой стороны, стоимость приспособленности данного потерянного обеда непредсказуема, так как хищник может быстро найти лучшую добычу. Кроме того, большинство хищников являются универсалами, что снижает влияние данной адаптации добычи на хищника. Поскольку специализация вызвана коэволюцией хищник-жертва, редкость специалистов может означать, что гонки вооружений хищник-жертва редки. [122]
Трудно определить, являются ли данные адаптации действительно результатом коэволюции, когда адаптация к добыче порождает адаптацию к хищнику, которая противостоит дальнейшей адаптации у добычи. Альтернативным объяснением является эскалация , когда хищники адаптируются к конкурентам, своим собственным хищникам или опасной добыче. [123] Очевидные адаптации к хищничеству могли также возникнуть по другим причинам, а затем быть кооптированы для нападения или защиты. У некоторых насекомых, на которых охотятся летучие мыши, слух развился до появления летучих мышей и использовался для того, чтобы слышать сигналы, используемые для территориальной защиты и спаривания. [124] Их слух развился в ответ на хищничество летучих мышей, но единственным явным примером взаимной адаптации у летучих мышей является скрытная эхолокация. [125]
Более симметричная гонка вооружений может происходить, когда добыча опасна, имеет шипы, иглы, токсины или яд, которые могут нанести вред хищнику. Хищник может ответить избеганием, что, в свою очередь, стимулирует эволюцию мимикрии. Избегание не обязательно является эволюционной реакцией, поскольку оно, как правило, усваивается из плохого опыта с добычей. Однако, когда добыча способна убить хищника (как это может сделать коралловая змея с ее ядом), нет возможности для обучения, и избегание должно быть унаследовано. Хищники также могут реагировать на опасную добычу контрадаптациями. В западной части Северной Америки обыкновенная подвязочная змея выработала устойчивость к токсину в коже тритона с грубой кожей . [122]
Хищники влияют на свои экосистемы не только напрямую, поедая свою собственную добычу, но и косвенными способами, такими как сокращение хищничества со стороны других видов или изменение поведения травоядных животных при поиске пищи, как в случае с влиянием волков на биоразнообразие прибрежной растительности или морских выдр на леса водорослей. Это может объяснить эффекты динамики популяции, такие как циклы, наблюдаемые у рысей и зайцев-беляков. [126] [127] [128]
Один из способов классификации хищников — по трофическому уровню . Плотоядные , которые питаются травоядными , являются вторичными потребителями; их хищники — третичными потребителями и т. д. [129] На вершине этой пищевой цепи находятся высшие хищники, такие как львы . [130] Однако многие хищники питаются с нескольких уровней пищевой цепи; плотоядное животное может есть как вторичных, так и третичных потребителей. [131] Это означает, что многим хищникам приходится бороться с внутригильдийным хищничеством , когда другие хищники убивают и поедают их. Например, койоты конкурируют с серыми лисицами и рысями , а иногда и убивают их . [132]
Трофический перенос в экосистеме относится к переносу энергии и питательных веществ в результате хищничества. Энергия переходит с одного трофического уровня на другой, когда хищники потребляют органические вещества из тела другого организма . В каждом переносе, хотя есть использование энергии, есть также и потери энергии.
Морские трофические уровни различаются в зависимости от местоположения и размера первичных производителей . Обычно в открытом океане существует до шести трофических уровней, четыре на континентальных шельфах и около трех в зонах апвеллинга. [133] Например, морскую среду обитания с пятью трофическими уровнями можно представить следующим образом: травоядные (питаются в основном фитопланктоном ); плотоядные (питаются в основном другим зоопланктоном / животными); детритофаги (питаются в основном мертвым органическим веществом / детритом ); всеядные (питаются смешанным рационом из фито- и зоопланктона и детрита); и миксотрофы , которые сочетают автотрофию (использование световой энергии для роста без потребления каких-либо дополнительных органических соединений или питательных веществ) с гетеротрофией (питание другими растениями и животными для получения энергии и питательных веществ — травоядные, всеядные и плотоядные, а также детритофаги).
Эффективность трофического переноса измеряет, насколько эффективно энергия передается или проходит через более высокие трофические уровни морской пищевой сети . По мере того, как энергия перемещается вверх по трофическим уровням, она уменьшается из-за тепла, отходов и естественных метаболических процессов , которые происходят, когда хищники потребляют свою добычу. В результате только около 10% энергии на любом трофическом уровне передается на следующий уровень. Это часто называют «правилом 10%», которое ограничивает количество трофических уровней, которые может поддерживать отдельная экосистема. [134]
Хищники могут увеличивать биоразнообразие сообществ, не давая одному виду стать доминирующим. Такие хищники известны как ключевые виды и могут оказывать глубокое влияние на баланс организмов в определенной экосистеме . [135] Внедрение или удаление этого хищника или изменение плотности его популяции может иметь радикальные каскадные эффекты на равновесие многих других популяций в экосистеме. Например, травоядные на лугу могут не допустить захвата власти одним доминирующим видом. [136]
_horiz.jpg/1280px-Fig._1_-Riparian_willow_recovery_(26485120926)_horiz.jpg)
Уничтожение волков в Йеллоустонском национальном парке оказало глубокое воздействие на трофическую пирамиду . В этой области волки являются как ключевым видом, так и высшими хищниками. Без хищничества травоядные начали чрезмерно выпасать многие виды древесных побегов, влияя на популяции растений в этом районе. Кроме того, волки часто не давали животным пастись вблизи ручьев, защищая источники пищи бобров . Уничтожение волков оказало прямое воздействие на популяцию бобров, поскольку их среда обитания стала территорией для выпаса. Увеличенное поедание ив и хвойных деревьев вдоль ручья Блэктейл из-за отсутствия хищников привело к прореживанию канала, поскольку сократившаяся популяция бобров больше не могла замедлять воду и удерживать почву на месте. Таким образом, было показано, что хищники имеют жизненно важное значение в экосистеме. [137]
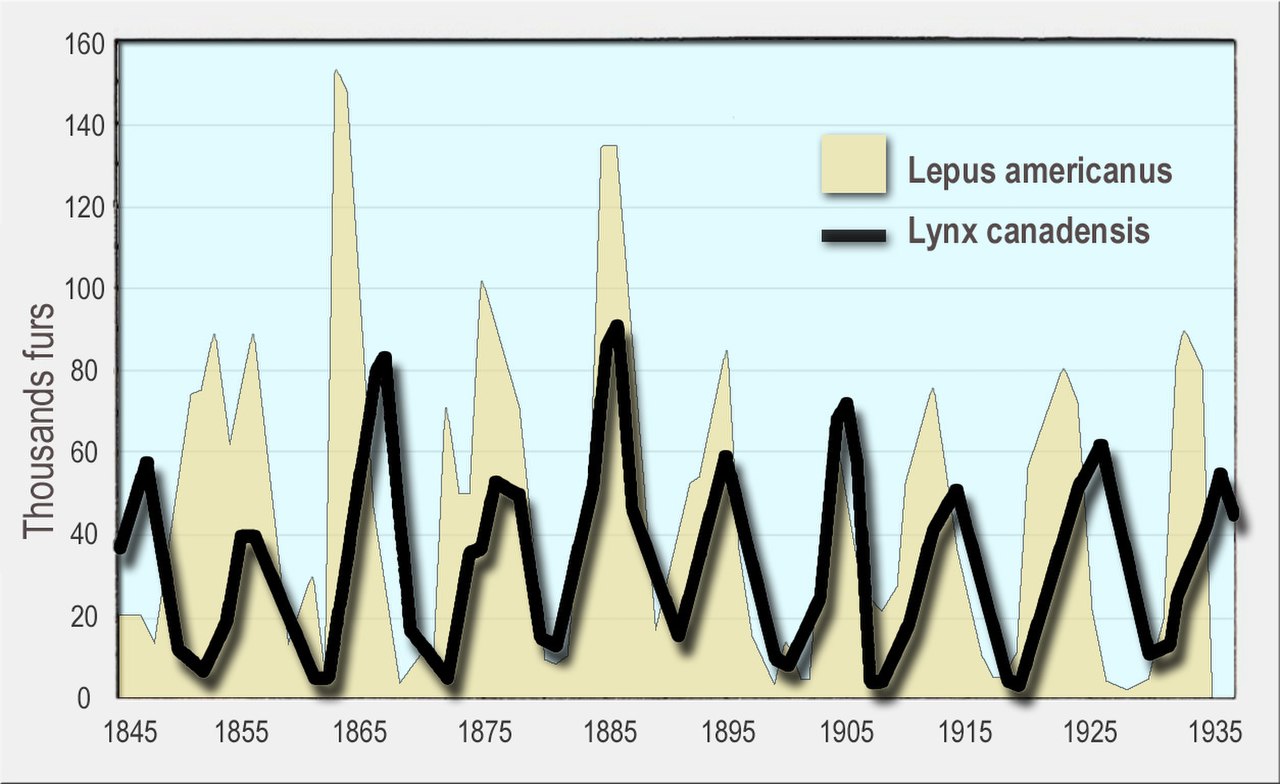
При отсутствии хищников популяция вида может расти экспоненциально, пока не достигнет предела емкости окружающей среды. [138] Хищники ограничивают рост добычи, как потребляя ее, так и изменяя ее поведение. [139] Увеличение или уменьшение популяции добычи может также привести к увеличению или уменьшению численности хищников, например, за счет увеличения числа детенышей, которых они рожают.
Циклические колебания наблюдались в популяциях хищников и жертв, часто со смещением между циклами хищников и жертв. Хорошо известным примером является заяц -беляк и рысь . На широком участке бореальных лесов на Аляске и в Канаде популяции зайцев колеблются почти синхронно с 10-летним периодом, а популяции рысей колеблются в ответ. Впервые это было замечено в исторических записях о животных, пойманных охотниками на пушного зверя для компании Hudson's Bay Company на протяжении более столетия. [140] [128] [141] [142]
.png/1280px-Lotka-Volterra_model_(1.1,_0.4,_0.4,_0.1).png)
Простая модель системы с одним видом хищника и одной жертвы, уравнения Лотки-Вольтерры , предсказывает популяционные циклы. [143] Однако попытки воспроизвести предсказания этой модели в лабораторных условиях часто терпели неудачу; например, когда простейшее Didinium nasutum добавляется к культуре, содержащей его добычу, Paramecium caudatum , последняя часто вымирает. [144]
Уравнения Лотки–Вольтерры опираются на несколько упрощающих предположений, и они структурно нестабильны , что означает, что любое изменение в уравнениях может стабилизировать или дестабилизировать динамику. [145] [146] Например, одно из предположений заключается в том, что хищники имеют линейную функциональную реакцию на добычу: скорость убийств увеличивается пропорционально скорости встреч. Если эта скорость ограничена временем, потраченным на обработку каждой добычи, то популяции добычи могут достичь плотности, выше которой хищники не смогут их контролировать. [144] Другое предположение заключается в том, что все особи добычи идентичны. В действительности хищники склонны выбирать молодых, слабых и больных особей, оставляя популяции добычи способными к восстановлению. [147]
Многие факторы могут стабилизировать популяции хищников и жертв. [148] Одним из примеров является наличие нескольких хищников, особенно универсалов, которых привлекает данный вид добычи, если он многочислен, и которые ищут его в другом месте, если его нет. [149] В результате циклы популяции, как правило, встречаются в северных умеренных и субарктических экосистемах, поскольку пищевые цепи там проще. [150] Система заяц-беляк-рысь является субарктической, но даже она включает в себя других хищников, включая койотов, ястребов-тетеревятников и больших рогатых сов , и цикл усиливается изменениями в пище, доступной зайцам. [151]
Ряд математических моделей был разработан путем ослабления предположений, сделанных в модели Лотки-Вольтерры; они по-разному позволяют животным иметь географическое распределение или мигрировать ; иметь различия между особями, такими как пол и возрастная структура , так что только некоторые особи размножаются; жить в изменяющейся среде, например, со сменой времен года ; [152] [153] и анализировать взаимодействия более чем двух видов одновременно. Такие модели предсказывают сильно различающуюся и часто хаотичную динамику популяции хищник-жертва. [152] [154] Наличие зон убежища , где добыча находится в безопасности от хищников, может позволить добыче поддерживать более крупные популяции, но также может дестабилизировать динамику. [155] [156] [157] [158]
Хищничество датируется временем до появления общепризнанных плотоядных животных на сотни миллионов (возможно, миллиарды) лет. Хищничество неоднократно развивалось в разных группах организмов. [5] [159] Рост эукариотических клеток около 2,7 млрд лет назад, рост многоклеточных организмов около 2 млрд лет назад и рост мобильных хищников (около 600 млн лет назад - 2 млрд лет назад, вероятно, около 1 млрд лет назад) - все это было приписано раннему хищническому поведению, и многие очень ранние останки показывают доказательства наличия скважин или других отметин, приписываемых мелким видам хищников. [5] Это, вероятно, вызвало основные эволюционные переходы, включая появление клеток , эукариот , полового размножения , многоклеточности , увеличения размера, подвижности (включая полет насекомых [160] ) и бронированных панцирей и экзоскелетов. [5]
Самыми ранними хищниками были микробные организмы, которые поглощали или поедали других. Поскольку ископаемые останки скудны, эти первые хищники могли датироваться где-то между 1 и более 2,7 Gya (миллиардами лет назад). [5] Хищничество стало заметно важным незадолго до кембрийского периода — около 550 миллионов лет назад — о чем свидетельствует почти одновременное развитие кальцификации у животных и водорослей, [161] и рытье нор, избегающее хищников . Однако хищники поедали микроорганизмы по крайней мере с 1000 миллионов лет назад , [5] [162] [163] с доказательствами избирательного (а не случайного) хищничества с того же времени. [164]
Auroralumina attenboroughii — эдиакарская кроновая группа книдарий (557–562 млн лет назад, около 20 млн лет до кембрийского взрыва) из леса Чарнвуд , Англия. Считается, что это одно из самых ранних хищных животных, ловящее мелкую добычу своими нематоцистами , как это делают современные книдарии. [165]
Ископаемые останки демонстрируют долгую историю взаимодействий между хищниками и их добычей, начиная с кембрийского периода, показывая, например, что некоторые хищники сверлили раковины двустворчатых и брюхоногих моллюсков, в то время как другие поедали эти организмы, разбивая их раковины. [166] Среди кембрийских хищников были беспозвоночные, такие как аномалокаридиды с придатками, подходящими для захвата добычи, большими сложными глазами и челюстями, сделанными из твердого материала, похожего на тот, что находится в экзоскелете насекомых. [167] Некоторые из первых рыб, у которых были челюсти, были бронированными и в основном хищными плакодермами силурийского и девонского периодов , один из которых , 6 - метровый (20 футов) дунклеостей , считается первым в мире позвоночным «суперхищником», охотящимся на других хищников. [168] [169] Насекомые развили способность летать в раннем карбоне или позднем девоне, что позволило им, среди прочего, спасаться от хищников. [160] Среди крупнейших хищников, которые когда-либо жили, были тероподовые динозавры, такие как тираннозавр из мелового периода. Они охотились на травоядных динозавров, таких как гадрозавры , цератопсы и анкилозавры . [170]


.png/1280px-Dunkleosteus_terrelli_(2024).png)

Люди, как всеядные , в некоторой степени являются хищниками, [171] используя оружие и инструменты для ловли рыбы , [172] охоты и ловли животных. [173] Они также используют другие виды хищников, такие как собаки , бакланы , [174] и соколы , чтобы ловить добычу для еды или ради спорта. [175] Два хищника среднего размера, собаки и кошки, являются животными, которых чаще всего держат в качестве домашних животных в западных обществах. [176] [177] Люди-охотники, включая сан из Южной Африки, используют охоту на упорство , форму преследования, при которой преследователь может быть медленнее добычи, такой как антилопа куду , на коротких дистанциях, но следует за ней в полуденную жару, пока она не истощится, преследование, которое может занять до пяти часов. [178] [179]
В биологической борьбе с вредителями хищники (и паразитоиды) из естественного ареала вредителя вводятся для контроля популяций, рискуя вызвать непредвиденные проблемы. Естественные хищники, при условии, что они не причиняют вреда видам, не являющимся вредителями, являются экологически чистым и устойчивым способом снижения ущерба посевам и альтернативой использованию химических агентов, таких как пестициды . [180]

В кино идея хищника как опасного гуманоидного врага используется в научно-фантастическом боевике ужасов 1987 года «Хищник» и его трех продолжениях . [181] [182] Ужасающий хищник, гигантский людоед | большая белая акула , пожирающая людей , также является центральным персонажем триллера Стивена Спилберга 1974 года «Челюсти » . [183]
Среди поэзии на тему хищничества можно исследовать сознание хищника, например, в стихотворении Теда Хьюза « Щука » . [184] Фраза «Природа, окровавленная зубами и когтями» из стихотворения Альфреда, лорда Теннисона 1849 года « In Memoriam AHH » была интерпретирована как относящаяся к борьбе между хищниками и добычей. [185]
В мифологии и народных сказках хищники, такие как лиса и волк, имеют неоднозначную репутацию. [186] Лиса была символом плодородия в Древней Греции, но погодным демоном в Северной Европе и созданием дьявола в раннем христианстве; лиса представлена как хитрая, жадная и хитрая в баснях, начиная с Эзопа . [186] Большой плохой волк известен детям по сказкам, таким как « Красная Шапочка» , но является демонической фигурой в исландских сагах «Эдда» , где волк Фенрир появляется в апокалиптическом конце света . [186] В Средние века распространилась вера в оборотней , людей, превращенных в волков. [186] В Древнем Риме и Древнем Египте волку поклонялись, волчица появляется в мифе об основании Рима, вскармливая Ромула и Рема . [186] Совсем недавно, в произведении Редьярда Киплинга «Книга джунглей» 1894 года , Маугли воспитывается стаей волков. [186] Отношение к крупным хищникам в Северной Америке, таким как волк, медведь гризли и пума, изменилось от враждебности или амбивалентности, сопровождавшейся активным преследованием, к позитивному и защитному во второй половине 20-го века. [187]
{{cite journal}}: Цитировать журнал требует |journal=( помощь )Стратегии беглой охоты варьируются от одной крайности кратковременного ускорения, мощности и скорости до другой крайности настойчивости и выносливости, при этом добыча утомляется для облегчения поимки. Считается, что собаки и люди полагаются на выносливость, а не на прямую скорость и маневренность для успеха при беглой охоте.
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка )ранние книги Хьюза содержали ошеломляющее изобилие стихотворений между обложками: ... рыбы и птицы, звери полевые и лесные, энергичные воплощения хищников и жертв. Будучи студентом, Хьюз занялся антропологией, а не литературой, и он решил медитировать, чтобы войти в трансоподобные состояния предсознания, прежде чем излагать стихи на бумаге. Его стихотворения, ранние или поздние, вступают в отношения живых существ; они приближаются к сознанию животных:
Мысль-лис,
Кот Эстер,
Щука.


.jpg/1280px-Hawk_eating_prey_(cropped).jpg)

![Auroralumina attenboroughii, эдиакарский хищник (ок. 560 млн лет назад). Это был книдарий стволовой группы, ловивший добычу нематоцистами.[165]](http://upload.wikimedia.org/wikipedia/commons/thumb/6/63/Auroralumina_attenboroughii_reconstruction.jpg/1280px-Auroralumina_attenboroughii_reconstruction.jpg)

