Млекопитающее (от лат. mamma 'грудь') [1] — позвоночное животное класса млекопитающих ( / m ə ˈ m eɪ l i . ə / ) . Млекопитающие характеризуются наличием молочных желез , вырабатывающих молоко для кормления детенышей, неокортексной области мозга, шерсти или волос и трех косточек среднего уха . Эти характеристики отличают их от рептилий и птиц , от которых их предки отделились в каменноугольном периоде более 300 миллионов лет назад. Описано около 6400 современных видов млекопитающих, разделенных на 29 отрядов .
Крупнейшими отрядами млекопитающих по числу видов являются грызуны , летучие мыши и эвлипотифлы (в том числе ежи , кроты и землеройки ). Следующие три — это приматы (включая людей , обезьян и лемуров ), парнокопытные (включая свиней , верблюдов и китов ) и хищные животные (включая кошек , собак и тюленей ).
Млекопитающие — единственные живые члены Synapsida ; эта клада вместе с Sauropsida (рептилиями и птицами) составляет более крупную кладу Amniota . Ранние синапсиды называются « пеликозаврами ». Более продвинутые терапсиды стали доминировать в средней перми . Млекопитающие произошли от цинодонтов , развитой группы терапсидов, в период от позднего триаса до ранней юры . Современные млекопитающие достигли своего современного разнообразия в палеогеновом и неогеновом периодах кайнозойской эры , после вымирания нептичьих динозавров , и были доминирующей группой наземных животных с 66 миллионов лет назад до настоящего времени.
Основной тип тела млекопитающих — четвероногий , и большинство млекопитающих используют свои четыре конечности для передвижения по земле ; но у некоторых конечности приспособлены к жизни на море , в воздухе , на деревьях , под землей или на двух ногах . Размер млекопитающих варьируется от шмелиной летучей мыши размером 30–40 мм (1,2–1,6 дюйма) до синего кита длиной 30 м (98 футов) — возможно, самого крупного животного, когда-либо жившего на свете. Максимальная продолжительность жизни варьируется от двух лет у землероек до 211 лет у гренландского кита . Все современные млекопитающие рождают живых детенышей, за исключением пяти видов однопроходных , которые являются яйцекладущими млекопитающими. Самая богатая видами группа млекопитающих, группа , называемая плацентами , имеет плаценту , которая обеспечивает питание плода во время беременности .
Большинство млекопитающих разумны , некоторые обладают большим мозгом, самосознанием и умением пользоваться инструментами . Млекопитающие могут общаться и издавать звуки несколькими способами, включая производство ультразвука , маркировку запахами , сигналы тревоги , пение , эхолокацию ; и, в случае с людьми, сложный язык . Млекопитающие могут организовываться в общества деления-слияния , гаремы и иерархии , но также могут быть одиночными и территориальными . Большинство млекопитающих полигинны , но некоторые могут быть моногамными или полиандрическими .
Одомашнивание многих видов млекопитающих людьми сыграло важную роль в неолитической революции и привело к тому, что сельское хозяйство заменило охоту и собирательство в качестве основного источника пищи для людей. Это привело к серьезной реструктуризации человеческого общества от кочевого к оседлому, с более тесным сотрудничеством между все большими и большими группами и, в конечном итоге, к развитию первых цивилизаций . Одомашненные млекопитающие обеспечивали и продолжают обеспечивать энергию для транспорта и сельского хозяйства, а также продукты питания ( мясо и молочные продукты ), мех и кожу . На млекопитающих также охотятся и гоняют в спортивных целях, и они используются в качестве модельных организмов в науке. Млекопитающие изображались в искусстве со времен палеолита и появляются в литературе, кино, мифологии и религии. Снижение численности и исчезновение многих млекопитающих вызваны, прежде всего, браконьерством и разрушением среды обитания , прежде всего вырубкой лесов .

Классификация млекопитающих претерпела несколько изменений с тех пор, как Карл Линней первоначально определил этот класс, и в настоящее время ни одна система классификации не является общепринятой. McKenna & Bell (1997) и Wilson & Reeder (2005) предоставили полезные недавние сборники. [2] Симпсон (1945) [3] представляет систематику происхождения и взаимоотношений млекопитающих, которой учили повсеместно до конца 20-го века. Однако с 1945 года постепенно было обнаружено большое количество новой и более подробной информации: палеонтологические данные были перекалиброваны, и за прошедшие годы наблюдалось много споров и прогресса относительно теоретических основ самой систематизации, отчасти благодаря новой концепции кладистика . Хотя полевые и лабораторные исследования постепенно устарели классификацию Симпсона, она остается наиболее близкой к официальной классификации млекопитающих, несмотря на известные проблемы. [4]
Большинство млекопитающих, включая шесть наиболее богатых видами отрядов , относятся к плацентарной группе. Три крупнейших по числу видов отряда — Rodentia : мыши , крысы , дикобразы , бобры , капибары и другие грызущие млекопитающие; Рукокрылые : летучие мыши; и Soricomorpha : землеройки , кроты и соленодоны . Следующие три крупнейших отряда, в зависимости от используемой схемы биологической классификации , — это приматы : человекообразные обезьяны , мартышки и лемуры ; Cetartiodactyla : киты и парнокопытные ; _ и Хищники , в которые входят кошки , собаки , ласки , медведи , тюлени и их союзники. [5] По данным Mammal Species of the World , в 2006 году было идентифицировано 5416 видов. Они были сгруппированы в 1229 родов , 153 семейства и 29 отрядов. [5] В 2008 году Международный союз охраны природы (МСОП) завершил пятилетнюю глобальную оценку млекопитающих для своего Красного списка МСОП , в котором насчитывалось 5488 видов. [6] Согласно исследованию, опубликованному в Журнале маммологии в 2018 году, количество признанных видов млекопитающих составляет 6495, включая 96 недавно вымерших. [7]
Слово «млекопитающее» является современным и происходит от научного названия Mammalia , придуманного Карлом Линнеем в 1758 году и происходящего от латинского mamma («соска, папочка»). В влиятельной статье 1988 года Тимоти Роу определил Mammalia филогенетически как коронную группу млекопитающих, кладу , состоящую из самого недавнего общего предка ныне живущих однопроходных ( ехидны и утконосы ) и терианских млекопитающих ( сумчатые и плацентарные ) и всех потомков этого предка. [8] Поскольку этот предок жил в юрском периоде, определение Роу исключает всех животных из более раннего триаса , несмотря на то, что триасовые окаменелости в харамиидах были отнесены к млекопитающим с середины 19 века. [9] Если Mammalia рассматривать как коронную группу, ее происхождение можно грубо датировать первым известным появлением животных, более тесно связанных с некоторыми современными млекопитающими, чем с другими. Амбондро более тесно связан с однопроходными, чем с териальными млекопитающими, в то время как амфилест и амфитерий более тесно связаны с терианами; поскольку окаменелости всех трех родов датируются примерно 167 миллионами лет назад в средней юре , это разумная оценка внешнего вида кроновой группы. [10]
Т. С. Кемп дал более традиционное определение: « Синапсиды , которые обладают зубно - чешуйчатым сочленением челюстей и окклюзией между верхними и нижними коренными зубами с поперечным компонентом движения» или, что то же самое, по мнению Кемпа, клада, происходящая от последнего общего предка Синоконодон и современные млекопитающие. [11] Самый ранний известный синапсид, удовлетворяющий определениям Кемпа, — это Tikitherium , датированный 225 млн лет назад , поэтому появление млекопитающих в этом более широком смысле можно отнести к этой дате позднего триаса . [12] [13]
В 1997 году млекопитающие были тщательно пересмотрены Малкольмом К. Маккенной и Сьюзен К. Белл, в результате чего была создана классификация Маккенна/Белл. Авторы вместе работали палеонтологами в Американском музее естественной истории . Маккенна унаследовал проект от Симпсона и вместе с Беллом построил полностью обновленную иерархическую систему, охватывающую живые и вымершие таксоны, которая отражает историческую генеалогию млекопитающих. [4] Их книга 1997 года « Классификация млекопитающих выше уровня вида » [14] представляет собой всеобъемлющую работу по систематике, взаимосвязям и распространению всех таксонов млекопитающих, ныне живущих и вымерших, вплоть до рода, хотя молекулярно-генетические данные подвергают сомнению несколько группировок.
В следующем списке вымершие группы отмечены кинжалом ( †).
Класс Млекопитающие

В начале 21 века молекулярные исследования, основанные на анализе ДНК , позволили предположить новые взаимоотношения между семействами млекопитающих. Большинство этих результатов были независимо подтверждены данными о присутствии/отсутствии ретротранспозонов . [17] Системы классификации, основанные на молекулярных исследованиях, выявляют три основные группы или линии плацентарных млекопитающих — Afrotheria , Xenarthra и Boreoeutheria , — которые разошлись в меловом периоде . Отношения между этими тремя линиями являются спорными, и были предложены все три возможные гипотезы относительно того, какая группа является базовой . Этими гипотезами являются Atlantogenata (базальная Boreoeutheria), Epitheria (базальная Xenarthra) и Exafroplacentalia (базальная Afrotheria). [18] Boreoeutheria, в свою очередь, включает две основные линии — Euarchontoglires и Laurasiatheria .
Оценки времени расхождения между этими тремя плацентарными группами варьируются от 105 до 120 миллионов лет назад, в зависимости от типа используемой ДНК (например, ядерной или митохондриальной ) [19] и различных интерпретаций палеогеографических данных. [18]
Synapsida , клада, в которую входят млекопитающие и их вымершие родственники, возникла во время пенсильванского субпериода (~323–300 миллионов лет назад), когда они отделились от линии рептилий. Млекопитающие кроновой группы произошли от более ранних млекопитающих в ранней юре . На кладограмме Mammalia считается кронной группой. [23]

Первыми полностью наземными позвоночными были амниоты . Как и их ранние четвероногие предшественники-амфибии, у них были легкие и конечности. Однако амниотические яйца имеют внутренние мембраны, которые позволяют развивающемуся эмбриону дышать, но удерживают воду. Следовательно, амниоты могут откладывать яйца на суше, тогда как земноводным обычно приходится откладывать яйца в воде.
Первые амниоты, по-видимому, возникли в пенсильванском подпериоде каменноугольного периода . Они произошли от более ранних четвероногих амфибий-рептилий, [24] которые жили на суше, уже заселенной насекомыми и другими беспозвоночными, а также папоротниками , мхами и другими растениями. В течение нескольких миллионов лет стали различаться две важные линии амниот: синапсиды , которые позже включали общего предка млекопитающих; и зауропсиды , к которым сейчас относятся черепахи , ящерицы , змеи , крокодилы и динозавры (включая птиц ). [25] Синапсиды имеют по одному отверстию ( височное окно ) внизу на каждой стороне черепа. Примитивные синапсиды включали самых крупных и свирепых животных ранней перми , таких как диметродон . [26] Синапсиды немлекопитающих традиционно (и неправильно) назывались «рептилиями, подобными млекопитающим» или пеликозаврами ; теперь мы знаем, что они не были ни рептилиями, ни частью линии рептилий. [27] [28]
Терапсиды , группа синапсид, развились в средней перми , около 265 миллионов лет назад, и стали доминирующими наземными позвоночными. [27] Они отличаются от базальных эвпеликозавров некоторыми особенностями черепа и челюстей, в том числе: более крупными черепами и резцами , которые у терапсидов одинаковы по размеру, но не у эвпеликозавров. [27] Линия терапсидов, ведущая к млекопитающим, прошла ряд стадий, начиная с животных, которые были очень похожи на своих ранних синапсидных предков, и заканчивая пробайногнатическими цинодонтами , некоторых из которых можно было легко принять за млекопитающих. Эти этапы характеризовались: [29]
Пермско -триасовое вымирание около 252 миллионов лет назад, которое было длительным событием из-за накопления нескольких импульсов вымирания, положило конец доминированию хищных терапсидов. [31] В раннем триасе большинство средних и крупных ниш наземных хищников были заняты архозаврами [32] , которые в течение длительного периода (35 миллионов лет) стали включать крокодиломорфов , [ 33] птерозавров и динозавров; [34] однако крупные цинодонты, такие как Trucidocynodon и traversodontids, все еще занимали крупные ниши плотоядных и травоядных соответственно. К юрскому периоду динозавры стали доминировать и в крупных нишах наземных травоядных. [35]
Первые млекопитающие (в понимании Кемпа) появились в эпоху позднего триаса (около 225 миллионов лет назад), через 40 миллионов лет после первых терапсидов. Начиная с середины юрского периода, они вышли из своей ночной ниши насекомоядных ; [36] Юрский касторокауда , например, был близким родственником настоящих млекопитающих, у которых были приспособления для плавания, рытья и ловли рыбы. [37] Считается, что большинство, если не все, продолжали вести ночной образ жизни ( ночное узкое место ), что объясняет большую часть типичных черт млекопитающих. [38] Большинство видов млекопитающих, существовавших в мезозойскую эру, были мультитуберкулятами, эвтриконодонтами и спалакотериидами . [39] Самый ранний известный метатерий — Sinodelphys , найденный в сланцах раннего мела возрастом 125 миллионов лет в северо-восточной китайской провинции Ляонин . Окаменелость почти полная и включает в себя пучки меха и отпечатки мягких тканей. [40]

Самая старая известная окаменелость среди Eutheria («настоящие звери») — это маленькая землеройка Juramaia sinensis , или «юрская мать из Китая», датируемая 160 миллионами лет назад, в позднем юрском периоде. [41] Более поздний родственник эвтерий, Eomaia , датируемый 125 миллионами лет назад в раннем меловом периоде, обладал некоторыми чертами, общими с сумчатыми, но не с плацентарными, что свидетельствует о том, что эти черты присутствовали у последнего общего предка двух групп. но позже были потеряны в плацентарной линии. [42] В частности, надлобковые кости выступают вперед от таза. Они не обнаружены ни в одной современной плаценте, но обнаружены у сумчатых, однопроходных, других нетерийных млекопитающих и Ukhaatherium , раннего мелового животного из эвтерийского отряда Asioryctitheria . Это относится и к мультитуберкулезам. [43] По-видимому, они являются наследственной особенностью, которая впоследствии исчезла в плацентарной линии. Эти надлобковые кости, по-видимому, функционируют, напрягая мышцы во время передвижения, уменьшая количество представляемого пространства, которое требуется плаценте для содержания плода во время беременности. Узкое выходное отверстие таза указывает на то, что детеныши при рождении были очень маленькими и поэтому беременность была короткой, как у современных сумчатых. Это говорит о том, что плацента возникла позднее. [44]
Одним из самых ранних известных однопроходных был тейнолофос , живший около 120 миллионов лет назад в Австралии. [45] Однопроходные имеют некоторые особенности, которые могут быть унаследованы от первоначальных амниот, например, одно и то же отверстие для мочеиспускания, дефекации и размножения ( клоака ) — как это делают ящерицы и птицы — [46] и они откладывают кожистые и некальцинированные яйца . [47]
Hadrocodium , чьи окаменелости датируются примерно 195 миллионами лет назад, в ранней юре , представляет собой первое четкое свидетельство существования челюстного сустава, образованного исключительно чешуйчатой и зубной костями; в челюсти нет места для суставной кости, входящей в состав челюстей всех ранних синапсид. [48]

Самые ранние явные свидетельства наличия волос или меха встречаются в окаменелостях Castorocauda и Megaconus , датированных 164 миллионами лет назад в середине юрского периода. В 1950-х годах было высказано предположение, что отверстия (ходы) в верхних челюстях и предчелюстных костях (костях передней части верхней челюсти) цинодонтов представляют собой каналы, которые снабжают кровеносные сосуды и нервы вибриссами ( усами ) и, таким образом, свидетельствуют о наличии волос или шерсть; [49] [50] однако вскоре было отмечено, что отверстия не обязательно указывают на то, что у животного были вибриссы, поскольку у современной ящерицы Tupinambis есть отверстия, которые почти идентичны тем, которые обнаружены у немлекопитающих цинодонтов Thrinaxodon . [28] [51] Популярные источники, тем не менее, продолжают приписывать усы тринаксодону . [52] Исследования пермских копролитов показывают, что синапсиды немлекопитающих той эпохи уже имели мех, что указывает на то, что эволюция волос, возможно, началась еще у дицинодонтов . [53]
Когда эндотермия впервые появилась в эволюции млекопитающих, точно неизвестно, хотя общепринято считать, что она впервые возникла у терапсидов , не относящихся к млекопитающим . [53] [54] Современные однопроходные имеют более низкую температуру тела и более изменчивую скорость метаболизма, чем сумчатые и плацентарные, [55] но есть свидетельства того, что некоторые из их предков, возможно, включая предков териев, могли иметь температуру тела, подобную температуре тела современные терианцы. [56] Точно так же у некоторых современных терианцев, таких как афротерии и ксенартраны, вторично развилась более низкая температура тела. [57]
Эволюция прямостоячих конечностей у млекопитающих не завершена: у живых и ископаемых однопроходных конечности раскидистые. Парасагиттальное (нераскинутое) положение конечностей появилось где-то в поздней юре или раннем мелу; он обнаружен у эвтерийных Eomaia и метатерийных Sinodelphys , возраст которых датируется 125 миллионами лет назад. [58] Надлобковые кости, особенность, которая сильно повлияла на размножение большинства клад млекопитающих, впервые обнаружены у Tritylodontidae , что позволяет предположить, что это синапоморфия между ними и млекопитающими . Они повсеместно присутствуют у неплацентарных млекопитающих, хотя у Megazostrodon и Erythrotherium они, по-видимому, отсутствуют. [59]
Было высказано предположение, что первоначальная функция лактации ( производства молока ) заключалась в поддержании яиц влажными. Большая часть аргументов основана на однопроходных, млекопитающих, откладывающих яйца. [60] [61] У женщин молочные железы полностью развиваются в период полового созревания, независимо от беременности. [62]

Терианские млекопитающие заняли средние и крупные экологические ниши в кайнозое , после того как мел-палеогеновое вымирание примерно 66 миллионов лет назад опустошило экологическое пространство, когда-то заполненное нептичьими динозаврами и другими группами рептилий, а также различными другими видами. группы млекопитающих, [64] и претерпели экспоненциальное увеличение размеров тела ( мегафауна ). [65] Затем млекопитающие очень быстро диверсифицировались; и птицы, и млекопитающие демонстрируют экспоненциальный рост разнообразия. [64] Например, самая ранняя известная летучая мышь датируется примерно 50 миллионами лет назад, всего через 16 миллионов лет после вымирания нептичьих динозавров. [66]
Молекулярно-филогенетические исследования первоначально предполагали, что большинство плацентарных отрядов разошлись примерно от 100 до 85 миллионов лет назад и что современные семьи появились в период с позднего эоцена по миоцен . [67] Однако никаких окаменелостей плаценты не было обнаружено до конца мелового периода. [68] Самые ранние бесспорные окаменелости плаценты относятся к раннему палеоцену , после вымирания нептичьих динозавров. [68] (Учёные идентифицировали животное раннего палеоцена по имени Protungulatum donnae как одно из первых плацентарных млекопитающих, [69] но с тех пор оно было реклассифицировано как неплацентарное плацентарное животное.) [70] Перекалибровка показателей генетического и морфологического разнообразия позволила предположить, что позднемеловое происхождение плаценты и палеоценовое происхождение большинства современных клад . [71]
Самым ранним известным предком приматов является Archicebus Achilles [72] , живший примерно 55 миллионов лет назад. [72] Этот крошечный примат весил 20–30 граммов (0,7–1,1 унции) и мог поместиться на человеческой ладони. [72]
Живые виды млекопитающих можно идентифицировать по наличию потовых желез , в том числе тех, которые специализируются на выработке молока для питания детенышей. [73] Однако при классификации окаменелостей необходимо использовать и другие признаки, поскольку в окаменелостях не видны железы мягких тканей и многие другие особенности. [74]
Многие черты, общие для всех ныне живущих млекопитающих, появились у самых ранних членов группы:
По большей части эти характеристики отсутствовали у триасовых предков млекопитающих. [80] Почти все млекопитающие обладают надлобковой костью, за исключением современных плацентарных. [81]

В среднем самцы млекопитающих крупнее самок, при этом самцы как минимум на 10% крупнее самок более чем у 45% исследованных видов. Большинство отрядов млекопитающих также демонстрируют половой диморфизм, ориентированный на самцов , хотя некоторые отряды не демонстрируют какой-либо предвзятости или имеют значительную предвзятость к самкам ( Lagomorpha ). Половой диморфизм размеров увеличивается с размером тела у млекопитающих ( правило Ренша ), что позволяет предположить, что существует параллельное давление отбора как на размеры самцов, так и самок. Диморфизм, ориентированный на самцов, относится к половому отбору самцов посредством конкуренции между самцами за самок, поскольку существует положительная корреляция между степенью полового отбора, на что указывают системы спаривания , и степенью диморфизма размера, ориентированного на самцов. Степень полового отбора также положительно коррелирует с размером самцов и самок млекопитающих. Кроме того, параллельное давление отбора на массу самок обнаруживается в том, что возраст отлучения от груди значительно выше у более полигинных видов, даже с поправкой на массу тела. Кроме того, скорость размножения ниже у более крупных самок, что указывает на то, что отбор по плодовитости у млекопитающих осуществляется в пользу более мелких самок. Хотя эти закономерности сохраняются для млекопитающих в целом, между отрядами существуют значительные различия. [82]
У большинства млекопитающих семь шейных позвонков (костей шеи). Исключением являются ламантин и двупалый ленивец , у которых их шесть, а также трехпалый ленивец, у которого их девять. [83] Мозг всех млекопитающих имеет неокортекс — область мозга, уникальную только для млекопитающих. [84] Плацентарный мозг имеет мозолистое тело , в отличие от однопроходных и сумчатых. [85]
Сердце млекопитающих состоит из четырех камер: двух верхних предсердий (приемных камер) и двух нижних желудочков (выбрасывающих камер). [86] Сердце имеет четыре клапана, которые разделяют его камеры и обеспечивают ток крови через сердце в правильном направлении (предотвращая обратный ток). После газообмена в легочных капиллярах (кровеносных сосудах легких) обогащенная кислородом кровь возвращается в левое предсердие через одну из четырех легочных вен . Кровь почти непрерывно течет обратно в предсердие, которое действует как приемная камера, и отсюда через отверстие в левый желудочек. Большая часть крови пассивно поступает в сердце, пока и предсердия, и желудочки расслаблены, но к концу периода релаксации желудочков левое предсердие сокращается, перекачивая кровь в желудочек. Сердцу также необходимы питательные вещества и кислород, которые содержатся в крови, как и другие мышцы, и снабжаются ими через коронарные артерии . [87]

Легкие млекопитающих губчатые и сотовые. Дыхание осуществляется преимущественно с помощью диафрагмы , которая отделяет грудную клетку от брюшной полости, образуя выпуклый к грудной клетке купол. Сокращение диафрагмы уплощает купол, увеличивая объем полости легкого. Воздух поступает через ротовую и носовую полости, проходит через гортань, трахею и бронхи , расширяет альвеолы . Расслабление диафрагмы имеет противоположный эффект: объем полости легких уменьшается, что приводит к вытеснению воздуха из легких. Во время тренировки брюшная стенка сокращается , увеличивая давление на диафрагму, которая выталкивает воздух быстрее и сильнее. Грудная клетка способна расширять и сжимать грудную полость под действием других дыхательных мышц. Следовательно, воздух всасывается в легкие или выбрасывается из них, всегда перемещаясь по градиенту давления. [88] [89] Этот тип легких известен как сильфонное легкое из-за его сходства с кузнечными сильфонами . [89]
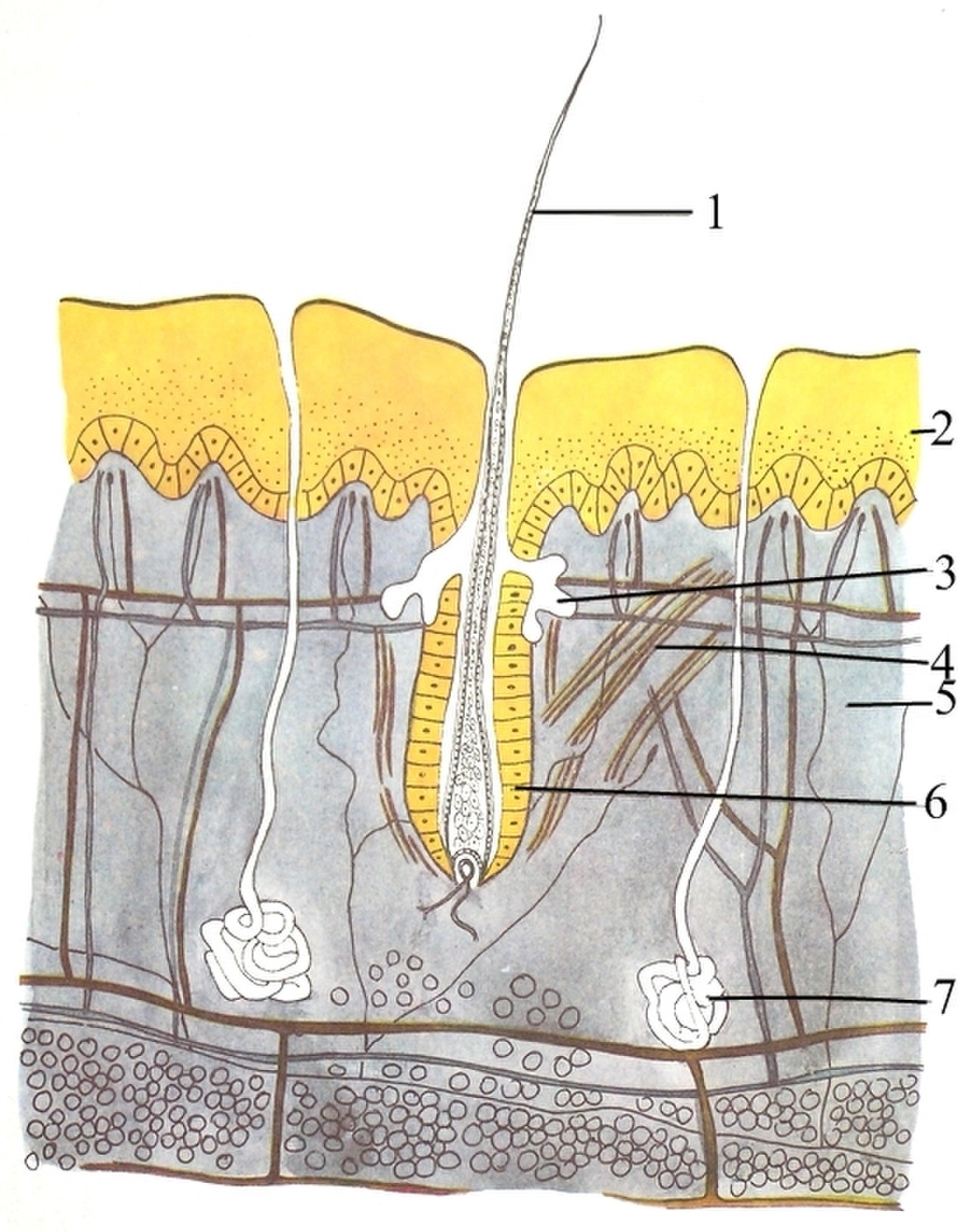
Покровная система (кожа) состоит из трех слоев: внешнего эпидермиса , дермы и гиподермы . Эпидермис обычно имеет толщину от 10 до 30 клеток; его основная функция — создание водонепроницаемого слоя. Его внешние клетки постоянно теряются; его самые нижние клетки постоянно делятся и продвигаются вверх. Средний слой, дерма, в 15–40 раз толще эпидермиса. Дерма состоит из многих компонентов, таких как костные структуры и кровеносные сосуды. Гиподерма состоит из жировой ткани , которая хранит липиды и обеспечивает амортизацию и изоляцию. Толщина этого слоя широко варьируется от вида к виду; [90] : 97 морским млекопитающим требуется толстая подкожа ( жир ) для изоляции, а у китов самый толстый жир — 20 дюймов (51 см). [91] Хотя у других животных есть такие особенности, как усы, перья , щетинки или реснички , которые внешне напоминают его, ни у одного животного, кроме млекопитающих, нет волос . Это определяющая характеристика класса, хотя у некоторых млекопитающих ее очень мало. [90] : 61
Травоядные животные развили разнообразные физические структуры, облегчающие потребление растительного материала . Чтобы разрушить неповрежденные растительные ткани, млекопитающие развили структуру зубов , отражающую их пищевые предпочтения. Например, плодоядные (животные, питающиеся в основном фруктами) и травоядные, питающиеся мягкой листвой, имеют зубы с низкой коронкой, предназначенные для измельчения листвы и семян . Пасущиеся животные, которые питаются твердой, богатой кремнеземом травой, имеют зубы с высокой коронкой, которые способны перемалывать жесткие ткани растений и не изнашиваются так быстро, как зубы с низкой коронкой. [92] У большинства плотоядных млекопитающих зубы карнассиальной формы (разной длины в зависимости от рациона), длинные клыки и схожие модели замещения зубов. [93]
Желудок парнокопытных (Artiodactyla) разделен на четыре отдела: рубец , сетку , книжку и сычуг (рубец есть только у жвачных ). После употребления растительного материала он смешивается со слюной в рубце и сетке и разделяется на твердый и жидкий материал. Твердые вещества комкаются вместе, образуя комок (или жвачку ), и отрыгиваются. При попадании комка в рот жидкость выдавливается языком и снова проглатывается. Поглощенная пища попадает в рубец и ретикулум, где целлюлолитические микробы ( бактерии , простейшие и грибы ) производят целлюлазу , которая необходима для расщепления целлюлозы в растениях. [94] Периссодактили , в отличие от жвачных животных, хранят переваренную пищу, покинувшую желудок, в увеличенной слепой кишке , где она ферментируется бактериями. [95] Хищные животные имеют простой желудок, приспособленный для переваривания преимущественно мяса, в отличие от сложной пищеварительной системы травоядных животных, которая необходима для расщепления жестких и сложных растительных волокон. Слепая кишка либо отсутствует, либо короткая и простая, а толстая кишка не мешотчатая или значительно шире тонкой кишки. [96]

_(20732795545).jpg/1280px-Image_from_page_702_of_%22Outlines_of_zoology%22_(1895)_(20732795545).jpg)
Выделительная система млекопитающих включает в себя множество компонентов. Как и большинство других наземных животных, млекопитающие являются уреотеликами и преобразуют аммиак в мочевину , что осуществляется печенью как часть цикла мочевины . [97] Билирубин , продукт жизнедеятельности клеток крови , выводится через желчь и мочу с помощью ферментов, выделяемых печенью. [98] Прохождение билирубина с желчью через кишечный тракт придает фекалиям млекопитающих характерную коричневую окраску. [99] К отличительным особенностям почек млекопитающих относится наличие почечной лоханки и почечных пирамид , а также четко различимых коркового и мозгового слоя , что обусловлено наличием удлиненных петель Генле . Только почка млекопитающих имеет бобовидную форму, хотя есть и некоторые исключения, например многолопастные ретикуляционные почки ластоногих, китообразных и медведей. [100] [101] У большинства взрослых плацентарных млекопитающих не осталось следов клоаки . У эмбриона эмбриональная клоака разделяется на задний отдел, который становится частью ануса, и передний отдел, судьба которого различна в зависимости от пола особи: у самок она развивается в преддверие или мочеполовой синус , в который попадает уретра. и влагалище , тогда как у мужчин он образует всю уретру полового члена . [101] [102] Однако тенреки , золотые кроты и некоторые землероек сохраняют клоаку и во взрослом возрасте. [103] У сумчатых половые пути отделены от ануса, но следы исходной клоаки все же остаются снаружи. [101] Monotremes, что переводится с греческого как «единственное отверстие», имеет настоящую клоаку. [104]
Как и у всех других четвероногих, у млекопитающих есть гортань , которая может быстро открываться и закрываться для воспроизведения звуков, а также надгортанный голосовой тракт , который фильтрует этот звук. Легкие и окружающая мускулатура обеспечивают поток воздуха и давление, необходимые для фонации . Гортань контролирует высоту и громкость звука, но сила, с которой легкие выдыхают, также способствует увеличению громкости. Более примитивные млекопитающие, например ехидна, могут только шипеть, поскольку звук достигается исключительно за счет выдоха через частично закрытую гортань. Другие млекопитающие голосуют с помощью голосовых связок . Движение или напряжение голосовых связок может привести к появлению множества звуков, таких как мурлыканье и крик . Млекопитающие могут менять положение гортани, позволяя им дышать через нос, глотать через рот, а также формировать как ротовые, так и носовые звуки; носовые звуки, такие как собачье вой, обычно тихие, а оральные звуки, такие как лай собаки, обычно громкие. [105]
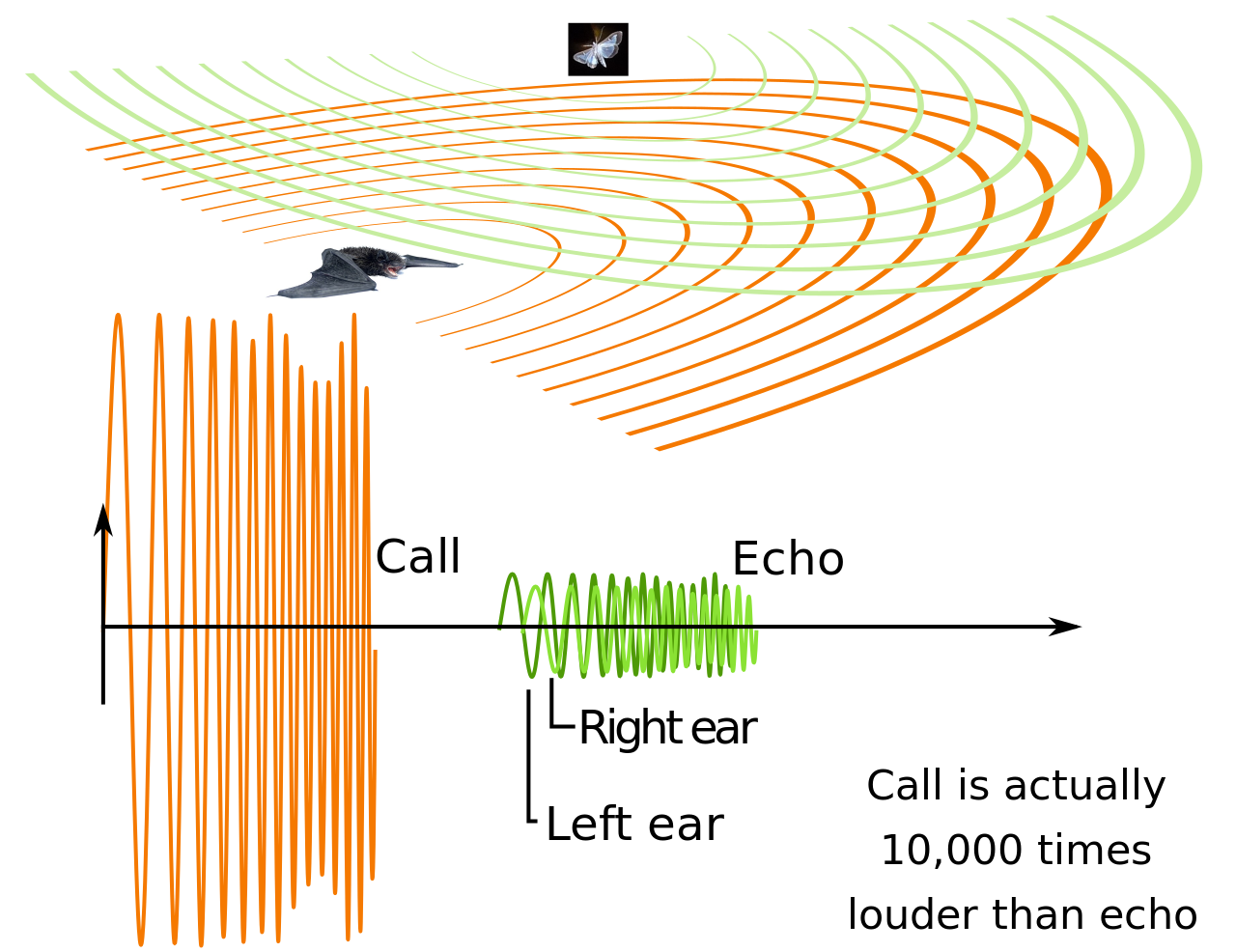
У некоторых млекопитающих большая гортань и, следовательно, низкий голос, а именно у летучей мыши-молота ( Hypsignathus monstrosus ), у которой гортань может занимать всю грудную полость , одновременно выталкивая легкие, сердце и трахею в брюшную полость . [106] Большие голосовые подушечки также могут понижать высоту звука, как в случае с низким ревом больших кошек . [107] Производство инфразвука возможно у некоторых млекопитающих, таких как африканский слон ( виды Loxodonta ) и усатые киты . [108] [109] Мелкие млекопитающие с маленькой гортанью обладают способностью производить ультразвук , который можно обнаружить путем модификаций среднего уха и улитки . Ультразвук не слышен птицам и рептилиям, что могло быть важно во времена мезозоя, когда птицы и рептилии были доминирующими хищниками. Этот частный канал используется некоторыми грызунами, например, при общении между матерью и щенком, а также летучими мышами при эхолокации. Зубатые киты также используют эхолокацию, но, в отличие от голосовой перепонки, простирающейся вверх от голосовых складок, у них есть дыня для манипулирования звуками. У некоторых млекопитающих, а именно приматов, к гортани прикреплены воздушные мешки, которые могут снижать резонансы или увеличивать громкость звука. [105]
Система голосовой продукции контролируется ядрами черепных нервов головного мозга и снабжается возвратным гортанным нервом и верхним гортанным нервом , ветвями блуждающего нерва . Голосовой тракт иннервируется подъязычным нервом и лицевыми нервами . Электрическая стимуляция околоводопроводной серой (ПЭГ) области среднего мозга млекопитающих вызывает вокализацию. Способность изучать новые вокализации наблюдается только у людей, тюленей, китообразных, слонов и, возможно, летучих мышей; у человека это результат прямой связи между моторной корой , управляющей движением, и мотонейронами спинного мозга. [105]

Основная функция меха млекопитающих — терморегуляция . Другие включают защиту, сенсорные цели, гидроизоляцию и камуфляж. [110] Разные виды меха служат разным целям: [90] : 99
Длина волос не является фактором терморегуляции: например, у некоторых тропических млекопитающих, таких как ленивцы, длина меха такая же, как у некоторых арктических млекопитающих, но с меньшей изоляцией; и, наоборот, другие тропические млекопитающие с короткой шерстью обладают такой же изолирующей способностью, как и арктические млекопитающие. Густота меха может повысить изоляционные свойства животного, а у арктических млекопитающих особенно густой мех; например, у овцебыка есть остевые волосы длиной 30 см (12 дюймов), а также густой подшерсток, образующий воздухонепроницаемую шерсть, позволяющую им выживать при температуре -40 ° C (-40 ° F). [90] : 162–163 Некоторые пустынные млекопитающие, такие как верблюды, используют густой мех, чтобы предотвратить попадание солнечного тепла на кожу, что позволяет животному сохранять прохладу; Летом температура верблюжьего меха может достигать 70 ° C (158 ° F), но температура кожи остается 40 ° C (104 ° F). [90] : 188 Водные млекопитающие , наоборот, задерживают воздух в своем мехе, чтобы сохранить тепло, сохраняя кожу сухой. [90] : 162–163

Шерсть млекопитающих окрашена по разным причинам, основные факторы отбора включают камуфляж , половой отбор , общение и терморегуляцию. Окраска как волос, так и кожи млекопитающих определяется главным образом типом и количеством меланина ; эумеланин для коричневого и черного цвета и феомеланин для диапазона от желтоватого до красноватого цвета, придающего млекопитающим землистый оттенок . [111] [112] Некоторые млекопитающие имеют более яркие цвета; некоторые обезьяны, такие как мандрилы и верветки , а также опоссумы, такие как мексиканские опоссумы-мыши и шерстистые опоссумы Дерби , имеют синюю кожу из-за дифракции света в коллагеновых волокнах. [113] Многие ленивцы кажутся зелеными, потому что в их мехе обитают зеленые водоросли ; это может быть симбиотическая связь, позволяющая ленивцам маскироваться . [114]
Камуфляж оказывает сильное влияние на большое количество млекопитающих, поскольку помогает скрыть особей от хищников или добычи. [115] У арктических и субарктических млекопитающих, таких как песец ( Alopex lagopus ), лемминг с ошейником ( Dicrostonyx groenlandicus ), горностай ( Mustela erminea ) и заяц-беляк ( Lepus americanus ), сезонное изменение цвета между коричневым летом и белым зимой. во многом обусловлено камуфляжем. [116] Некоторые древесные млекопитающие, особенно приматы и сумчатые, имеют оттенки фиолетового, зеленого или синего цвета кожи на частях тела, что указывает на определенное преимущество в их преимущественно древесной среде обитания благодаря конвергентной эволюции . [113]
Апосематизм , предупреждающий возможных хищников, является наиболее вероятным объяснением черно-белой шерсти многих млекопитающих, способных защитить себя, например, у дурно пахнущего скунса и мощного и агрессивного медоеда . [117] Цвет шерсти иногда имеет половой диморфизм , как у многих видов приматов . [118] Различия в цвете шерсти самок и самцов могут указывать на питание и уровень гормонов, что важно при выборе партнера. [119] Цвет шерсти может влиять на способность сохранять тепло в зависимости от того, сколько света отражается. Млекопитающие с более темной шерстью могут поглощать больше тепла от солнечной радиации и оставаться теплее, а у некоторых более мелких млекопитающих, таких как полевки , зимой мех темнее. Белый беспигментированный мех арктических млекопитающих, таких как белый медведь, может отражать больше солнечной радиации непосредственно на кожу. [90] : 166–167 [110] Ослепительные черно-белые полосы зебр , по-видимому, обеспечивают некоторую защиту от укусов мух. [120]

Млекопитающие исключительно гонохорны (животное рождается либо с мужскими, либо с женскими гениталиями, в отличие от гермафродитов , у которых такого раскола нет). [121] В мужских плацентах пенис используется как для мочеиспускания, так и для совокупления. В зависимости от вида эрекция может вызываться притоком крови в сосуды, губчатую ткань или мышечным действием. В не эрегированном состоянии половой член может находиться в крайней плоти , а в некоторых плацентах также имеется кость полового члена ( бакулюм ). [122] Сумчатые обычно имеют раздвоенные пенисы, [123] тогда как пенис ехидны обычно имеет четыре головки, из которых функционируют только две. [124] У большинства млекопитающих яички опускаются в мошонку , которая обычно находится позади полового члена, но у сумчатых часто находится спереди. У самок млекопитающих обычно имеется вульва ( клитор и половые губы ) снаружи, тогда как внутренняя система содержит парные яйцеводы , 1–2 матки , 1–2 шейки матки и влагалище . [125] [126] Сумчатые имеют два боковых влагалища и медиальное влагалище. «Влагалище» однопроходных лучше понимать как «мочеполовой синус». Маточные системы плацентарных млекопитающих могут различаться между дуплексной, когда имеются две матки и шейки матки, открывающиеся во влагалище, двудольной, когда два рога матки имеют одну шейку, которая соединяется с влагалищем, и двурогой , состоящей из двух маточных систем. рога, которые соединены дистально, но разделены медиально, образуя Y-образную форму, и симплекс, имеющий единственную матку. [127] [128] [90] : 220–221, 247

Исходным условием размножения млекопитающих является рождение относительно неразвитых особей либо посредством прямого живорождения , либо в течение короткого периода в виде яиц с мягкой скорлупой. Вероятно, это связано с тем, что туловище не могло расширяться из-за наличия надлобковых костей . Самая старая демонстрация этого репродуктивного стиля - Kayentatherium , который произвел неразвитых перинатов , но в помете гораздо большего размера, чем у любого современного млекопитающего - 38 экземпляров. [129] Большинство современных млекопитающих являются живородящими и рождают живых детенышей. Однако пять видов однопроходных, утконос и четыре вида ехидны откладывают яйца. У однопроходных система определения пола отличается от системы большинства других млекопитающих. [130] В частности, половые хромосомы утконоса больше похожи на хромосомы курицы, чем на хромосомы терийского млекопитающего. [131]
Живородящие млекопитающие относятся к подклассу Theria; живущие сегодня относятся к сумчатым и плацентарным инфраклассам. У сумчатых период беременности короткий , обычно короче эстрального цикла , и обычно они рождают несколько неразвитых новорожденных, которые затем подвергаются дальнейшему развитию; у многих видов это происходит внутри мешочка, сумчатого , расположенного в передней части живота матери . Это плезиоморфное состояние среди живородящих млекопитающих; наличие надлобковых костей у всех неплацентарных млекопитающих препятствует расширению туловища, необходимому для полноценной беременности. [81] Даже неплацентарные особи, вероятно, размножались таким образом. [43] Плаценты рождают относительно полноценных и развитых детенышей, обычно после длительных периодов беременности. [132] Свое название они получили от плаценты , которая соединяет развивающийся плод со стенкой матки, обеспечивая поглощение питательных веществ. [133] У плацентарных млекопитающих надлобковая кость либо полностью утрачена, либо преобразована в бакулюм; позволяя туловищу расширяться и, таким образом, рождать развитое потомство. [129]
Молочные железы млекопитающих специализируются на выработке молока, основного источника питания новорожденных. Однопроходные рано ответвились от других млекопитающих и не имеют сосков, которые наблюдаются у большинства млекопитающих, но у них есть молочные железы. Детеныши слизывают молоко с участка молочной железы на животе матери. [134] По сравнению с плацентарными млекопитающими, молоко сумчатых сильно меняется как по производительности, так и по составу питательных веществ из-за недоразвитости молодняка. Кроме того, молочные железы обладают большей автономией, что позволяет им поставлять отдельное молоко детенышам на разных стадиях развития. [135] Лактоза является основным сахаром в молоке плацентарных млекопитающих, тогда как в молоке однопроходных и сумчатых преобладают олигосахариды . [136] Отлучение от груди — это процесс, при котором млекопитающее становится менее зависимым от материнского молока и больше от твердой пищи. [137]
Почти все млекопитающие эндотермичны («теплокровны»). У большинства млекопитающих также есть волосы, которые помогают им сохранять тепло. Как и птицы, млекопитающие могут добывать пищу или охотиться в погоде и климате, слишком холодных для экзотермических («хладнокровных») рептилий и насекомых. Эндотермия требует большого количества пищевой энергии, поэтому млекопитающие съедают больше пищи на единицу массы тела, чем большинство рептилий. [138] Мелкие насекомоядные млекопитающие едят огромные количества для своего размера. Редкое исключение — голый землекоп выделяет мало метаболического тепла, поэтому его считают оперативным пойкилотермом . [139] Птицы также эндотермичны, поэтому эндотермия не уникальна для млекопитающих. [140]
Среди млекопитающих максимальная продолжительность жизни видов значительно варьируется (например, продолжительность жизни землеройки составляет два года, тогда как зарегистрированный возраст самого старого гренландского кита составляет 211 лет). [141] Хотя основная причина этих различий в продолжительности жизни все еще неясна, многочисленные исследования показывают, что способность восстанавливать повреждения ДНК является важным фактором, определяющим продолжительность жизни млекопитающих. В исследовании 1974 года Харта и Сетлоу [142] было обнаружено, что способность к эксцизионному восстановлению ДНК систематически увеличивается с увеличением продолжительности жизни семи видов млекопитающих. Было обнаружено, что продолжительность жизни видов тесно коррелирует со способностью распознавать двухцепочечные разрывы ДНК, а также с уровнем белка репарации ДНК Ku80 . [141] В исследовании клеток шестнадцати видов млекопитающих было обнаружено, что гены, участвующие в репарации ДНК, активируются у долгоживущих видов. [143] При исследовании 13 видов млекопитающих было обнаружено, что клеточный уровень фермента репарации ДНК поли-АДФ-рибозо-полимеразы коррелирует с продолжительностью жизни видов. [144] Три дополнительных исследования различных видов млекопитающих также сообщили о корреляции между продолжительностью жизни видов и способностью к восстановлению ДНК. [145] [146] [147]

Большинство позвоночных животных — земноводные, рептилии и некоторые млекопитающие, такие как человек и медведи — стопоходящие , ходят на всей нижней стороне стопы. Многие млекопитающие, такие как кошки и собаки, ходят на пальцах ног, причем большая длина шага обеспечивает большую скорость. Некоторые животные , например лошади , негулиградны и ходят на кончиках пальцев. Это еще больше увеличивает длину шага и, следовательно, скорость. [148] Известно также, что некоторые млекопитающие, а именно человекообразные обезьяны, ходят на костяшках пальцев , по крайней мере, на передних ногах. Гигантские муравьеды [149] и утконосы [150] также умеют ходить на костяшках пальцев. Некоторые млекопитающие являются двуногими и используют для передвижения только две конечности, что можно увидеть, например, у людей и человекообразных обезьян. Двуногие виды имеют большее поле зрения, чем четвероногие, сохраняют больше энергии и способны манипулировать объектами руками, что помогает в поиске пищи. Вместо того, чтобы идти, некоторые двуногие прыгают, например, кенгуру и кенгуру-крысы . [151] [152]
Животные будут использовать разные походки в зависимости от скорости, местности и ситуаций. Например, лошади демонстрируют четыре естественных аллюра: самый медленный аллюр — это шаг , затем есть три более быстрых аллюра, от самого медленного к самому быстрому: рысь , галоп и галоп . Животные также могут иметь необычную походку, которую иногда используют, например, для движения вбок или назад. Например, основными человеческими походками являются ходьба и бег на двух ногах, но иногда они используют и многие другие походки, включая ползание на четырех ногах в ограниченном пространстве. [153] Млекопитающие демонстрируют широкий диапазон походки , порядок, в котором они размещают и поднимают свои придатки при передвижении. Походки можно сгруппировать по категориям в соответствии с их последовательностью поддержки. Четвероногих животных можно разделить на три основные категории: ходьба, бег и прыжки . [154] Ходьба — это наиболее распространенная походка, при которой некоторые ступни в любой момент времени стоят на земле, и встречается почти у всех четвероногих животных. Считается, что бег происходит, когда в некоторые моменты шага все ступни отрываются от земли в момент зависания. [153]
.jpg/1280px-Brachiating_Gibbon_(Some_rights_reserved).jpg)
Древесные животные часто имеют удлиненные конечности, которые помогают им преодолевать промежутки, добираться до фруктов или других ресурсов, проверять твердость опоры впереди и, в некоторых случаях, раскачиваться ( качаться между деревьями). [155] Многие древесные виды, такие как древесные дикобразы, шелковистые муравьеды , обезьяны-пауки и опоссумы , используют цепкие хвосты , чтобы хвататься за ветки. У обезьяны-паука кончик хвоста имеет либо оголенный участок, либо липкую подушечку, обеспечивающую повышенное трение. Когти можно использовать для взаимодействия с грубым субстратом и изменения направления сил, прилагаемых животным. Именно это позволяет белкам взбираться на стволы деревьев, которые настолько велики, что с точки зрения такого маленького животного они кажутся практически плоскими. Однако когти могут помешать животному хватать очень маленькие ветки, поскольку они могут слишком сильно обхватить и уколоть собственную лапу животного. Приматы используют фрикционный захват, полагаясь на безволосые кончики пальцев. Сжатие ветки между кончиками пальцев создает силу трения, которая удерживает руку животного на ветке. Однако этот тип захвата зависит от угла силы трения, то есть от диаметра ветки, причем более крупные ветви приводят к снижению способности захвата. Чтобы контролировать спуск, особенно по ветвям большого диаметра, у некоторых древесных животных, таких как белки, развились очень подвижные голеностопные суставы, которые позволяют поворачивать ступню в «перевернутое» положение. Это позволяет когтям цепляться за шероховатую поверхность коры, противодействуя силе тяжести. Небольшой размер дает много преимуществ древесным видам: например, увеличение относительного размера ветвей для животного, более низкий центр массы, повышенную устойчивость, меньшую массу (что позволяет передвигаться по более мелким ветвям) и способность перемещаться по более загроможденной среде обитания. [155] Размер, связанный с весом, влияет на планирующих животных, таких как сахарный планер . [156] Некоторые виды приматов, летучих мышей и все виды ленивцев достигают пассивной устойчивости, зависая под веткой. И подача, и опрокидывание становятся неуместными, поскольку единственным способом провала будет потеря хватки. [155]
Летучие мыши — единственные млекопитающие, которые по-настоящему умеют летать. Они летают по воздуху с постоянной скоростью, двигая крыльями вверх и вниз (обычно также с некоторым движением вперед-назад). Поскольку животное находится в движении, относительно его тела существует некоторый поток воздуха, который в сочетании со скоростью крыльев создает более быстрый поток воздуха, движущийся по крылу. Это создает вектор подъемной силы, направленный вперед и вверх, и вектор силы сопротивления, направленный назад и вверх. Направленные вверх компоненты противодействуют гравитации, удерживая тело в воздухе, в то время как передний компонент обеспечивает тягу, противодействующую сопротивлению как крыла, так и тела в целом. [157]
Крылья летучих мышей намного тоньше и состоят из большего количества костей, чем у птиц, что позволяет летучим мышам более точно маневрировать и летать с большей подъемной силой и меньшим сопротивлением. [158] [159] Складывая крылья внутрь по направлению к телу при взмахе вверх, они используют на 35% меньше энергии во время полета, чем птицы. [160] Оболочки нежные, легко рвутся; однако ткань мембраны летучей мыши способна отрастать заново, поэтому небольшие разрывы быстро заживают. [161] Поверхность их крыльев оснащена сенсорными рецепторами на небольших выпуклостях, называемых клетками Меркеля , которые также встречаются на кончиках пальцев человека. Эти чувствительные области у летучих мышей различны, так как каждая шишка имеет крошечный волосок в центре, что делает ее еще более чувствительной и позволяет летучей мыши обнаруживать и собирать информацию о воздухе, проходящем через ее крылья, а также летать более эффективно, изменяя форму. своих крыльев в ответ. [162]
Ископаемое (от латинского fossor , что означает «копатель») — животное, приспособленное к рытью, которое живет в основном, но не исключительно, под землей. Некоторые примеры — барсуки и голые землекопы . Многие виды грызунов также считаются ископаемыми, поскольку они живут в норах большую часть дня, но не весь день. Виды, живущие исключительно под землей, являются подземными, а виды с ограниченной адаптацией к роющему образу жизни — субфоссорными. Некоторые организмы являются ископаемыми, чтобы помочь в регулировании температуры , в то время как другие используют подземную среду обитания для защиты от хищников или для хранения пищи . [163]
Ископаемые млекопитающие имеют веретенообразное тело, наиболее толстое в плечах и суживающееся к хвосту и носу. Неспособные видеть в темных норах, у большинства деградировали глаза, но дегенерация варьируется у разных видов; Карманные суслики , например, имеют лишь полуфоссорный вид и имеют очень маленькие, но функциональные глаза, у полностью окаменелого сумчатого крота глаза деградировали и бесполезны, у тальповых кротов рудиментарные глаза, а у плащового золотого крота глаза покрыты слоем кожи. . Наружные ушные раковины также очень малы или отсутствуют. У истинно окаменелых млекопитающих короткие и толстые ноги, поскольку для роющих млекопитающих сила важнее скорости, а у полуфоссорных млекопитающих ноги бегающие . Передние лапы широкие и имеют сильные когти, которые помогают разрыхлять землю при рытье нор, а на задних лапах есть перепонки и когти, которые помогают отбрасывать разрыхленную землю назад. У большинства из них большие резцы, чтобы грязь не попадала в рот. [164]
Многие ископаемые млекопитающие, такие как землеройки, ежи и кроты, были отнесены к ныне устаревшему отряду Insectivora . [165]
Полностью водные млекопитающие, китообразные и сирены , лишились ног и имеют хвостовой плавник, позволяющий передвигаться по воде. Движение ласт непрерывное. Киты плавают, перемещая хвостовой плавник и нижнюю часть тела вверх и вниз, двигаясь вертикально, а их ласты в основном используются для управления. Их скелетная анатомия позволяет им быть быстрыми пловцами. У большинства видов есть спинной плавник , который не позволяет им переворачиваться в воде. [166] [167] Лапы сирен поднимаются вверх и вниз длинными движениями, чтобы переместить животное вперед, и могут поворачиваться для поворота. Передние конечности представляют собой ласты, похожие на весла, которые помогают поворачиваться и замедляться. [168]
Полуводные млекопитающие, такие как ластоногие, имеют две пары ласт спереди и сзади: передние и задние. Локти и лодыжки заключены внутри тела. [169] [170] Ластоногие имеют несколько приспособлений для уменьшения сопротивления . В дополнение к обтекаемому телу, у них есть гладкая сеть мышечных пучков на коже, которая может увеличивать ламинарный поток и облегчать им скольжение в воде. У них также отсутствуют выпрямляющие пили , поэтому их мех может быть обтекаемым во время плавания. [171] Для передвижения они полагаются на передние ласты, подобно крыльям, подобно пингвинам и морским черепахам . [172] Движение передних ласт не является непрерывным, и животное скользит между каждым гребком. [170] По сравнению с наземными хищниками, передние конечности уменьшены в длине, что дает локомоторным мышцам в плечевых и локтевых суставах большее механическое преимущество; [169] Задние ласты служат стабилизаторами. [171] Другие полуводные млекопитающие включают бобров, гиппопотамов , выдр и утконосов. [173] Бегемоты — очень крупные полуводные млекопитающие, их бочкообразные тела имеют гравипортальные скелетные структуры, [174] приспособленные к переносу их огромного веса, а их удельный вес позволяет им тонуть и перемещаться по дну реки. [175]

Многие млекопитающие общаются посредством вокала. Голосовое общение служит многим целям, в том числе в брачных ритуалах, в качестве предупреждающих сигналов , [177] для указания источников пищи, а также для социальных целей. Самцы часто кричат во время брачных ритуалов, чтобы отогнать других самцов и привлечь самок, например, рев львов и благородных оленей . [178] Песни горбатого кита могут быть сигналами для самок ; [179] В разных регионах океана у них разные диалекты. [180] Социальные вокализации включают территориальные крики гиббонов и использование частоты у больших остроносых летучих мышей для различения групп. [181] Верветка подает отчетливый сигнал тревоги для каждого из как минимум четырех различных хищников, и реакции других обезьян различаются в зависимости от сигнала. Например, если сигнал тревоги сигнализирует питону, обезьяны забираются на деревья, тогда как сигнал тревоги орла заставляет обезьян искать укрытие на земле. [176] Луговые собачки также издают сложные крики, которые сигнализируют о типе, размере и скорости приближающегося хищника. [182] Слоны социально общаются с помощью различных звуков, включая фырканье, крик, трубный рев, рев и урчание. Некоторые из грохочущих криков являются инфразвуковыми , ниже диапазона слышимости человека, и их могут услышать другие слоны на расстоянии до 6 миль (9,7 км) в неподвижное время перед восходом и закатом солнца. [183]
Млекопитающие подают сигналы различными способами. Многие подают визуальные сигналы против хищников , например, когда олени и газели останавливаются , честно указывая на свою физическую форму и способность убежать, [184] [185] или когда белохвостый олень и другие млекопитающие-жертвы сигнализируют с заметными отметинами на хвосте, когда встревожены, информируя хищника о том, что он обнаружен. [186] Многие млекопитающие используют запаховую маркировку , иногда, возможно, для защиты территории, но, вероятно, с рядом функций как внутри вида, так и между видами. [187] [188] [189] Летучие мыши и зубатые киты , включая океанических дельфинов, издают звуки как социально, так и с помощью эхолокации . [190] [191] [192]
Поддержание высокой постоянной температуры тела требует больших затрат энергии, поэтому млекопитающие нуждаются в питательном и обильном рационе. Хотя самые ранние млекопитающие, вероятно, были хищниками, с тех пор различные виды адаптировались к удовлетворению своих пищевых потребностей различными способами. Некоторые едят других животных — это плотоядная диета (включая насекомоядную). Другие млекопитающие, называемые травоядными , питаются растениями, которые содержат сложные углеводы, такие как целлюлоза. Травоядная диета включает такие подтипы, как зерноядные (поедание семян), фоливорные (поедание листьев), плодоядные (поедание фруктов), нектариворные (поедание нектара), мармеладные (поедание жевательной резинки) и микофагия (поедание грибов). Пищеварительный тракт травоядных животных является домом для бактерий, которые ферментируют эти сложные вещества и делают их доступными для пищеварения, которые находятся либо в многокамерном желудке , либо в большой слепой кишке. [94] Некоторые млекопитающие являются копрофагами и потребляют фекалии для поглощения питательных веществ, не переваренных при первом приеме пищи. [90] : 131–137 Всеядное животное питается как добычей, так и растениями. У плотоядных млекопитающих простой пищеварительный тракт , поскольку белки , липиды и минералы, содержащиеся в мясе, практически не требуют специального пищеварения. Исключением являются усатые киты , у которых кишечная флора также находится в многокамерном желудке, как и у наземных травоядных. [193]
Размер животного также является фактором, определяющим тип диеты ( правило Аллена ). Поскольку у мелких млекопитающих высокое соотношение площади теплоотдающей поверхности к теплогенерирующему объему, у них, как правило, высокие потребности в энергии и высокая скорость метаболизма . Млекопитающие весом менее 18 унций (510 г; 1,1 фунта) в основном насекомоядны, поскольку не переносят медленный и сложный процесс пищеварения травоядных. С другой стороны, более крупные животные выделяют больше тепла и теряют меньше этого тепла. Поэтому они могут переносить либо более медленный процесс сбора (хищники, питающиеся более крупными позвоночными), либо более медленный процесс пищеварения (травоядные). [194] Кроме того, млекопитающие, которые весят более 18 унций (510 г; 1,1 фунта), обычно не могут собрать достаточно насекомых в часы бодрствования, чтобы поддерживать себя. Единственными крупными насекомоядными млекопитающими являются те, которые питаются огромными колониями насекомых ( муравьев или термитов ). [195]
Некоторые млекопитающие всеядны и демонстрируют различную степень плотоядности и травоядности, обычно склоняясь к одному больше, чем к другому. Поскольку растения и мясо перевариваются по-разному, одно предпочтение отдается другому, как у медведей, у которых некоторые виды могут быть в основном плотоядными, а другие - преимущественно травоядными. [197] Они сгруппированы в три категории: мезокарниворные (50–70% мяса), гиперплотоядные (70% и более мяса) и гипохищные (50% и менее мяса). Зубной ряд гипоплотоядных животных состоит из тупых треугольных хищных зубов, предназначенных для перемалывания пищи. Гиперплотоядные животные, однако, имеют конические зубы и острые хищники, предназначенные для рубящих ударов, а в некоторых случаях сильные челюсти для дробления костей, как в случае с гиенами , что позволяет им поедать кости; у некоторых вымерших групп, особенно Machairodontinae , были клыки саблевидной формы . [196]
Некоторые физиологические хищники потребляют растительные вещества, а некоторые физиологические травоядные животные потребляют мясо. С поведенческой точки зрения это сделало бы их всеядными, но с физиологической точки зрения это может быть связано с зоофармакогнозией . Физиологически, чтобы считаться всеядными, животные должны иметь возможность получать энергию и питательные вещества из растительных и животных материалов. Таким образом, таких животных по-прежнему можно классифицировать как плотоядных и травоядных, хотя они только получают питательные вещества из материалов, происходящих из источников, которые, казалось бы, не дополняют их классификацию. [198] Например, хорошо известно, что некоторые копытные, такие как жирафы, верблюды и крупный рогатый скот, грызут кости, чтобы потреблять определенные минералы и питательные вещества. [199] Кроме того, кошки, которых обычно считают облигатными хищниками, иногда едят траву, чтобы отрыгнуть неперевариваемый материал (например, комки шерсти ), помочь в выработке гемоглобина и в качестве слабительного. [200]
Многие млекопитающие при отсутствии достаточной потребности в пище в окружающей среде подавляют свой метаболизм и сохраняют энергию в процессе, известном как гибернация . [201] В период, предшествующий спячке, более крупные млекопитающие, такие как медведи, становятся полифагами , чтобы увеличить запасы жира, тогда как более мелкие млекопитающие предпочитают собирать и хранить пищу. [202] Замедление метаболизма сопровождается снижением частоты сердечных сокращений и дыхания, а также падением внутренней температуры, которая в некоторых случаях может приближаться к температуре окружающей среды. Например, внутренняя температура зимующих арктических сусликов может упасть до -2,9 ° C (26,8 ° F); однако температура головы и шеи всегда остается выше 0 ° C (32 ° F). [203] Некоторые млекопитающие в жарких регионах впадают в спячку во время засухи или сильной жары, например, толстохвостый карликовый лемур ( Chirogaleus medius ). [204]
У разумных млекопитающих, таких как приматы , головной мозг больше по сравнению с остальной частью мозга. Интеллект сам по себе определить нелегко, но к признакам интеллекта относится способность к обучению, сочетающаяся с поведенческой гибкостью. Крысы , например, считаются очень умными, поскольку они могут учиться и выполнять новые задачи, и эта способность может быть важна, когда они впервые колонизируют новую среду обитания . У некоторых млекопитающих сбор пищи, по-видимому, связан с интеллектом: у оленя, питающегося растениями, мозг меньше, чем у кошки, и ей приходится думать, чтобы перехитрить свою добычу. [195]

Использование инструментов животными может указывать на разные уровни обучения и познания . Морская выдра использует камни как неотъемлемую и регулярную часть своего добывающего поведения (разбивая морское ушко о камни или разбивая раковины), при этом некоторые популяции тратят 21% своего времени на изготовление инструментов. [205] Другое использование инструментов, например, когда шимпанзе используют ветки для «ловли» термитов, может быть развито путем наблюдения за тем, как другие используют инструменты , и может даже быть истинным примером обучения животных. [206] Инструменты можно даже использовать при решении головоломок, в которых животное, кажется, переживает «момент Эврики» . [207] Другие млекопитающие, которые не используют инструменты, например собаки, также могут пережить момент Эврики. [208]
Размер мозга ранее считался основным показателем интеллекта животного. Поскольку большая часть мозга используется для поддержания функций организма, большее соотношение массы мозга к массе тела может увеличить количество массы мозга, доступной для более сложных когнитивных задач. Аллометрический анализ показывает, что размер мозга млекопитающих составляет примерно 2 ⁄ 3 или 3 ⁄ 4 показателя массы тела. Сравнение размера мозга конкретного животного с ожидаемым размером мозга на основе такого аллометрического анализа дает коэффициент энцефализации , который можно использовать как еще один показатель интеллекта животного. [209] Кашалоты имеют самую большую массу мозга среди всех животных на Земле: в среднем она составляет 8000 кубических сантиметров (490 куб. дюймов) и 7,8 кг (17 фунтов) у половозрелых самцов. [210]
Самосознание представляется признаком абстрактного мышления. Самосознание, хотя и не имеет четкого определения, считается предшественником более продвинутых процессов, таких как метакогнитивное мышление . Традиционным методом измерения этого показателя является зеркальный тест , который определяет, обладает ли животное способностью самопознания. [211] К млекопитающим, прошедшим зеркальный тест, относятся азиатские слоны (некоторые проходят, некоторые нет); [212] шимпанзе; [213] бонобо ; [214] орангутанги ; [215] человек, от 18 месяцев ( зеркальная стадия ); [216] обыкновенные афалины ; [а] [217] косатки ; [218] и ложные косатки . [218]

Эусоциальность – это высший уровень социальной организации. В этих обществах наблюдается перекрытие взрослых поколений, разделение репродуктивного труда и совместная забота о молодёжи. Обычно насекомые, такие как пчелы , муравьи и термиты, обладают эусоциальным поведением, но оно продемонстрировано у двух видов грызунов: голого землекопа [219] и дамаралендского землекопа . [220]
Пресоциальность — это когда животные демонстрируют больше, чем просто сексуальные взаимодействия с представителями того же вида, но не соответствуют критериям эусоциальности. То есть досоциальные животные могут демонстрировать коллективный образ жизни, совместную заботу о молодняке или примитивное разделение репродуктивного труда, но они не проявляют всех трех основных черт эусоциальных животных. Человек и некоторые виды Callitrichidae ( игрунки и тамарины ) уникальны среди приматов по степени совместной заботы о потомстве. [221] Гарри Харлоу поставил эксперимент на макаках-резусах , досоциальных приматах, в 1958 году; Результаты этого исследования показали, что социальные контакты необходимы для развития молодых обезьян как умственно, так и сексуально. [222]
Общество разделения-слияния — это общество, которое часто меняет свой размер и состав, образуя постоянную социальную группу, называемую «родительской группой». Постоянные социальные сети состоят из всех отдельных членов сообщества и часто меняются, чтобы отслеживать изменения в их среде. В обществе разделения-слияния основная родительская группа может распадаться (расщепляться) на более мелкие стабильные подгруппы или отдельных лиц, чтобы адаптироваться к экологическим или социальным обстоятельствам. Например, несколько самцов могут отделиться от основной группы, чтобы охотиться или добывать пищу в течение дня, но ночью они могут вернуться, чтобы присоединиться (слиться) с основной группой, чтобы поделиться едой и принять участие в других занятиях. Это наблюдается у многих млекопитающих, таких как приматы (например, орангутанги и паукообразные обезьяны ), [223] слоны, [224] пятнистые гиены , [225] львы, [226] и дельфины. [227]
Одиночные животные защищают территорию и избегают социальных взаимодействий с представителями своего вида, за исключением периода размножения. Это сделано для того, чтобы избежать конкуренции за ресурсы, поскольку две особи одного и того же вида будут занимать одну и ту же нишу, а также для предотвращения истощения запасов пищи. [228] Одиночное животное во время кормления также может быть менее заметным для хищников или добычи. [229]

В иерархии люди либо доминируют, либо подчиняются. Деспотическая иерархия — это когда одна особь доминирует, а другие подчиняются, как у волков и лемуров, [230] а иерархия — это линейное ранжирование особей, в котором есть высшая особь и низшая особь. Иерархия также может быть ранжирована по полу: самая низкая особь одного пола имеет более высокий рейтинг, чем высшая особь другого пола, как у гиен. [231] Доминирующие особи, или альфы, имеют высокие шансы на репродуктивный успех, особенно в гаремах , где один или несколько самцов (постоянных самцов) имеют исключительные права на размножение самок в группе. [232] Иногородние самцы также могут быть приняты в гаремы, но некоторые виды, такие как обыкновенная летучая мышь-вампир ( Desmodus rotundus ), могут быть более строгими. [233]
Некоторые млекопитающие совершенно моногамны , то есть они спариваются на всю жизнь и не принимают других партнеров (даже после смерти первоначального партнера), как волки, евразийские бобры и выдры. [234] [235] Существует три типа полигамии: один или несколько доминирующих самцов имеют право на размножение ( полигиния ), несколько самцов, с которыми спариваются самки (полиандрия), или несколько самцов имеют исключительные отношения с несколькими самками ( полигинандрия ). Гораздо чаще случается полигинное спаривание, которое, за исключением токов , по оценкам, встречается у 90% млекопитающих. [236] Спаривание леков происходит, когда самцы собираются вокруг самок и пытаются привлечь их различными проявлениями ухаживания и вокалами, как у обыкновенных тюленей. [237]
У всех высших млекопитающих (за исключением однопроходных) есть два основных приспособления для ухода за детенышами: живорождение и лактация. Они подразумевают общегрупповой выбор степени родительской заботы . Они могут строить гнезда и рыть норы, чтобы выращивать в них птенцов, или часто кормить и охранять их в течение длительного периода времени. Многие млекопитающие прошли K-отбор и вкладывают больше времени и энергии в развитие своего потомства, чем животные , отобранные r . Когда два животных спариваются, они оба заинтересованы в успехе потомства, хотя часто в разных крайностях. Самки млекопитающих проявляют некоторую степень материнской агрессии, что является еще одним примером родительской заботы, которая может быть направлена против других самок этого вида или детенышей других самок; однако некоторые млекопитающие могут «теть» детенышей других самок и заботиться о них. Самцы млекопитающих могут играть роль в воспитании детей, как и в случае с тенреками, однако это варьируется от вида к виду, даже в пределах одного рода. Например, самцы южной косичьей макаки ( Macaca nemestrina ) не участвуют в уходе за детьми, тогда как самцы японской макаки ( M. fuscata ) участвуют. [238]

Нечеловеческие млекопитающие играют самые разнообразные роли в человеческой культуре. Они являются самыми популярными домашними животными : десятки миллионов собак, кошек и других животных, включая кроликов и мышей, содержатся семьями по всему миру. [239] [240] [241] Млекопитающие, такие как мамонты , лошади и олени, являются одними из самых ранних предметов искусства, их можно найти в наскальных рисунках верхнего палеолита , таких как в Ласко . [242] Крупнейшие художники, такие как Альбрехт Дюрер , Джордж Стаббс и Эдвин Ландсир , известны своими портретами млекопитающих. [243] На многие виды млекопитающих охотились ради спорта и ради еды; Олени и кабаны особенно популярны в качестве охотничьих животных . [244] [245] [246] Млекопитающие, такие как лошади и собаки , широко участвуют в спортивных гонках, часто в сочетании со ставками на результат . [247] [248] Существует противоречие между ролью животных как товарищей по отношению к человеку и их существованием как личностей с собственными правами . [249] Млекопитающие также играют самые разнообразные роли в литературе, [250] [251] [252] кино, [253] мифологии и религии. [254] [255] [256]

Одомашнивание млекопитающих сыграло важную роль в развитии сельского хозяйства и цивилизации в эпоху неолита , заставив фермеров заменить охотников-собирателей по всему миру. [b] [258] Этот переход от охоты и собирательства к выпасу стад и выращиванию сельскохозяйственных культур был важным шагом в истории человечества. Новая сельскохозяйственная экономика, основанная на одомашненных млекопитающих, вызвала «радикальную реструктуризацию человеческого общества, глобальные изменения в биоразнообразии и значительные изменения в формах рельефа Земли и ее атмосфере... важные результаты». [259]
Домашние млекопитающие составляют большую часть поголовья скота , выращиваемого на мясо во всем мире. В их число входят (2009 г.) около 1,4 млрд голов крупного рогатого скота , 1 млрд овец , 1 млрд домашних свиней , [260] [261] и (1985 г.) более 700 млн кроликов. [262] Рабочие домашние животные , включая крупный рогатый скот и лошадей, использовались для работы и транспорта с самого начала сельского хозяйства, их численность снижается с появлением механизированного транспорта и сельскохозяйственной техники . В 2004 году они по-прежнему обеспечивали около 80% электроэнергии преимущественно мелких ферм в странах третьего мира и около 20% мирового транспорта, опять же в основном в сельских районах. В горных районах, непригодных для колесного транспорта, грузы продолжают перевозить вьючные животные. [263] Из шкур млекопитающих делают кожу для обуви , одежды и обивки . Шерсть млекопитающих, включая овец, коз и альпак , веками использовалась для изготовления одежды. [264] [265]

Млекопитающие играют важную роль в науке в качестве экспериментальных животных , как в фундаментальных биологических исследованиях, таких как генетика, [267] , так и в разработке новых лекарств, которые должны быть тщательно проверены, чтобы продемонстрировать их безопасность . [268] Ежегодно в экспериментах используются миллионы млекопитающих, особенно мышей и крыс . [269] Нокаутная мышь представляет собой генетически модифицированную мышь с инактивированным геном , замененным или разрушенным искусственным участком ДНК. Они позволяют изучать секвенированные гены, функции которых неизвестны. [270] Небольшой процент млекопитающих — это приматы, не относящиеся к человеку, которые используются в исследованиях из-за их сходства с людьми. [271] [272] [273]
Несмотря на пользу, которую одомашненные млекопитающие принесли для развития человечества, люди оказывают все более пагубное воздействие на диких млекопитающих во всем мире. Подсчитано, что масса всех диких млекопитающих сократилась до 4% от всех млекопитающих, при этом 96% млекопитающих сейчас составляют люди и их домашний скот (см. рисунок). Фактически наземные дикие млекопитающие составляют лишь 2% всех млекопитающих. [274] [275]
Гибриды — это потомство, полученное в результате скрещивания двух генетически различных особей, что обычно приводит к высокой степени гетерозиготности, хотя гибрид и гетерозигота не являются синонимами. Преднамеренная или случайная гибридизация двух или более видов близкородственных животных посредством разведения в неволе — это человеческая деятельность, которая существует на протяжении тысячелетий и развивается в экономических целях. [276] Гибриды между различными подвидами внутри вида (например, между бенгальским тигром и сибирским тигром ) известны как внутривидовые гибриды. Гибриды между разными видами одного рода (например, между львами и тиграми) известны как межвидовые гибриды или скрещивания. Гибриды между разными родами (например, между овцами и козами) известны как межродовые гибриды. [277] Естественные гибриды будут возникать в гибридных зонах , где две популяции видов одного и того же рода или виды, живущие в одних и тех же или соседних областях, будут скрещиваться друг с другом. Некоторые гибриды были признаны видами, например красный волк (хотя это спорный вопрос). [278]
Искусственный отбор , преднамеренное селекционное разведение домашних животных, используется для восстановления недавно вымерших животных в попытке создать породу животных с фенотипом , напоминающим вымершего предка дикого типа . Воскресный (внутривидовой) гибрид может быть очень похож на вымерший дикий тип по внешнему виду, экологической нише и в некоторой степени генетике, но исходный генофонд этого дикого типа навсегда утрачен с его исчезновением . В результате выведенные породы являются в лучшем случае расплывчатыми двойниками вымерших диких типов, так же как крупный рогатый скот Хека имеет сходство с зубром . [279]
Чистокровным диким видам, эволюционировавшим до специфической экологии, может угрожать исчезновение [280] из-за процесса генетического загрязнения , неконтролируемой гибридизации, интрогрессивного генетического затопления , которое приводит к гомогенизации или вытеснению гетерозисных гибридных видов. [281] Когда новые популяции импортируются или селективно разводятся людьми, или когда изменение среды обитания приводит к контакту ранее изолированных видов, возможно исчезновение некоторых видов, особенно редких разновидностей. [282] Скрещивание может поглотить более редкий генофонд и создать гибриды, истощая чистопородный генофонд. Например, находящийся под угрозой исчезновения дикий водяной буйвол находится под наибольшей угрозой исчезновения из-за генетического загрязнения домашним водяным буйволом . Такие вымирания не всегда очевидны с морфологической точки зрения. Некоторая степень потока генов является нормальным эволюционным процессом, тем не менее, гибридизация ставит под угрозу существование редких видов. [283] [284]

Утрата видов из экологических сообществ, дефаунация , в первую очередь обусловлена деятельностью человека. [285] Это привело к появлению пустых лесов , экологических сообществ, лишенных крупных позвоночных. [286] [287] Во время четвертичного вымирания массовое вымирание представителей мегафауны совпало с появлением людей, что свидетельствует о человеческом влиянии. Одна из гипотез заключается в том, что люди охотились на крупных млекопитающих, таких как шерстистый мамонт , и привели к их исчезновению. [288] [289] В докладе МПБЭУ о глобальной оценке биоразнообразия и экосистемных услуг за 2019 год говорится, что общая биомасса диких млекопитающих сократилась на 82 процента с момента возникновения человеческой цивилизации . [290] [291] Дикие животные составляют всего 4% биомассы млекопитающих на Земле, тогда как люди и их одомашненные животные составляют 96%. [275]
По прогнозам , в ближайшем будущем различные виды вымрут , [292] среди них носороги , [293] жирафы , [294] и виды приматов [295] и ящеры . [296] Согласно отчету WWF « Живая планета» за 2020 год , популяции позвоночных диких животных сократились на 68% с 1970 года в результате деятельности человека, особенно чрезмерного потребления , роста населения и интенсивного земледелия , что является свидетельством того, что люди спровоцировали шестое массовое вымирание . . [297] [298] Одна только охота угрожает сотням видов млекопитающих по всему миру. [299] [300] Ученые утверждают, что растущий спрос на мясо способствует утрате биоразнообразия , поскольку это является важным фактором вырубки лесов и разрушения среды обитания ; богатые видами места обитания, такие как значительная часть тропических лесов Амазонки , превращаются в сельскохозяйственные угодья для производства мяса. [301] [302] [303] Другим фактором влияния является чрезмерная охота и браконьерство , которые могут сократить общую популяцию охотничьих животных, [304] особенно тех, которые расположены вблизи деревень, [305] как в случае с пекари . [306] Последствия браконьерства особенно заметны в торговле слоновой костью африканских слонов. [307] Морские млекопитающие, особенно китообразные , подвергаются риску запутывания в рыболовных снастях , при этом смертность от выброса колеблется от 65 000 до 86 000 особей в год. [308]
Внимание уделяется исчезающим видам во всем мире, в частности, через Конвенцию о биологическом разнообразии , также известную как Соглашение Рио-де-Жанейро, в которую входят 189 подписавших стран, которые сосредоточены на выявлении исчезающих видов и мест обитания. [309] Еще одной известной природоохранной организацией является МСОП, в состав которого входят более 1200 правительственных и неправительственных организаций. [310]
Недавние вымирания можно напрямую объяснить влиянием человека. [311] [285] МСОП характеризует «недавнее» вымирание как те, которые произошли после порогового значения в 1500 лет, [312] и около 80 видов млекопитающих вымерли с этого времени и в 2015 году . [313] Некоторые виды, такие как олень Папа Давида [314] вымерли в дикой природе и выживают исключительно в популяциях, содержащихся в неволе. Другие виды, такие как пантера Флориды , экологически вымерли , выживая в таких небольших количествах, что практически не оказывают никакого влияния на экосистему. [315] : 318 Другие популяции вымерли лишь локально (искоренены), все еще существуют в других местах, но их распространение сократилось, [315] : 75–77, как и в случае с вымиранием серых китов в Атлантике . [316]
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на январь 2024 г. ( ссылка ){kind=link}