
Биология — это научное изучение жизни . [1] [2] [3] Это естественная наука с широким охватом, но имеющая несколько объединяющих тем, которые связывают ее в единую, связную область. [1] [2] [3] Например, все организмы состоят из клеток , которые обрабатывают наследственную информацию, закодированную в генах , которая может передаваться будущим поколениям. Другая важная тема — эволюция , которая объясняет единство и разнообразие жизни. [1] [2] [3] Обработка энергии также важна для жизни, поскольку она позволяет организмам двигаться , расти и размножаться . [1] [2] [3] Наконец, все организмы способны регулировать свою собственную внутреннюю среду . [1] [2] [3] [4] [5]
Биологи способны изучать жизнь на разных уровнях организации , [1] от молекулярной биологии клетки до анатомии и физиологии растений и животных, а также эволюции популяций. [1] [6] Следовательно, в биологии существует множество субдисциплин , каждая из которых определяется характером своих исследовательских вопросов и инструментами , которые они используют. [7] [8] [9] Как и другие ученые, биологи используют научный метод для проведения наблюдений , постановки вопросов, выдвижения гипотез , проведения экспериментов и формирования выводов об окружающем мире. [1]
Жизнь на Земле, возникшая более 3,7 миллиардов лет назад, [10] чрезвычайно разнообразна. Биологи стремились изучать и классифицировать различные формы жизни, от прокариотических организмов, таких как археи и бактерии, до эукариотических организмов, таких как протисты, грибы, растения и животные. Эти различные организмы вносят вклад в биоразнообразие экосистемы , где они играют специализированные роли в круговороте питательных веществ и энергии через свою биофизическую среду .

Самые ранние корни науки, включая медицину, можно проследить до Древнего Египта и Месопотамии примерно с 3000 по 1200 год до н. э . [11] [12] Их вклад сформировал древнегреческую натурфилософию . [13] [11] [12] [14] [15] Древнегреческие философы, такие как Аристотель (384–322 гг. до н. э.), внесли большой вклад в развитие биологических знаний. Он исследовал биологическую причинность и разнообразие жизни. Его преемник, Теофраст , начал научное изучение растений. [16] Ученые средневекового исламского мира , писавшие о биологии, включали аль-Джахиза (781–869), ад-Динавари (828–896), писавшего о ботанике, [17] и Разеса (865–925), писавшего об анатомии и физиологии . Медицина особенно хорошо изучалась исламскими учеными, работавшими в традициях греческой философии, в то время как естественная история во многом опиралась на философию Аристотеля.
Биология начала быстро развиваться с резким усовершенствованием микроскопа Антоном ван Левенгуком . Именно тогда ученые открыли сперматозоиды , бактерии, инфузории и разнообразие микроскопической жизни. Исследования Яна Сваммердама привели к новому интересу к энтомологии и помогли разработать методы микроскопического препарирования и окрашивания . [18] Достижения в области микроскопии оказали глубокое влияние на биологическое мышление. В начале 19 века биологи указали на центральное значение клетки . В 1838 году Шлейден и Шванн начали продвигать теперь уже универсальные идеи о том, что (1) основной единицей организмов является клетка и (2) что отдельные клетки обладают всеми характеристиками жизни, хотя они выступали против идеи о том, что (3) все клетки происходят из деления других клеток, продолжая поддерживать спонтанное зарождение . Однако Роберт Ремак и Рудольф Вирхов смогли обосновать третий принцип, и к 1860-м годам большинство биологов приняли все три принципа, которые объединились в клеточную теорию . [19] [20]
Между тем, таксономия и классификация стали центром внимания естествоиспытателей. Карл Линней опубликовал базовую таксономию для естественного мира в 1735 году, а в 1750-х годах ввел научные названия для всех своих видов. [21] Жорж-Луи Леклерк, граф де Бюффон , рассматривал виды как искусственные категории, а жизненные формы — как податливые, даже предполагая возможность общего происхождения . [22]

Серьёзное эволюционное мышление зародилось в работах Жана-Батиста Ламарка , который представил последовательную теорию эволюции. [24] Британский натуралист Чарльз Дарвин , объединив биогеографический подход Гумбольдта , униформистскую геологию Лайелла , труды Мальтуса о росте населения и свой собственный морфологический опыт и обширные естественные наблюдения, создал более успешную эволюционную теорию, основанную на естественном отборе ; схожие рассуждения и доказательства привели Альфреда Рассела Уоллеса к независимому приходу к тем же выводам. [25] [26]
Основа современной генетики началась с работы Грегора Менделя в 1865 году. [27] В ней были изложены принципы биологического наследования. [28] Однако значимость его работы не была осознана до начала 20-го века, когда эволюция стала единой теорией, поскольку современный синтез примирил дарвиновскую эволюцию с классической генетикой . [29] В 1940-х и начале 1950-х годов серия экспериментов Альфреда Херши и Марты Чейз указала на ДНК как на компонент хромосом , содержащий несущие признаки единицы, которые стали известны как гены . Сосредоточение внимания на новых видах модельных организмов, таких как вирусы и бактерии, наряду с открытием двойной спиральной структуры ДНК Джеймсом Уотсоном и Фрэнсисом Криком в 1953 году, ознаменовало переход к эре молекулярной генетики . С 1950-х годов биология значительно расширилась в молекулярной области. Генетический код был взломан Хар Гобиндом Кораной , Робертом В. Холли и Маршаллом Уорреном Ниренбергом после того, как стало известно, что ДНК содержит кодоны . Проект «Геном человека» был запущен в 1990 году для картирования генома человека . [30]
Все организмы состоят из химических элементов ; [31] кислород , углерод , водород и азот составляют большую часть (96%) массы всех организмов, а кальций , фосфор , сера , натрий , хлор и магний составляют по сути все остальное. Различные элементы могут объединяться, образуя соединения , такие как вода, которая имеет основополагающее значение для жизни. [31] Биохимия — это изучение химических процессов внутри живых организмов и связанных с ними . Молекулярная биология — это раздел биологии, который стремится понять молекулярную основу биологической активности в клетках и между ними, включая молекулярный синтез, модификацию, механизмы и взаимодействия.

Жизнь возникла из первого океана Земли, который образовался около 3,8 миллиарда лет назад. [32] С тех пор вода продолжает оставаться самой распространенной молекулой в каждом организме. Вода важна для жизни, потому что она является эффективным растворителем , способным растворять растворенные вещества, такие как ионы натрия и хлорида или другие небольшие молекулы, образуя водный раствор . После растворения в воде эти растворенные вещества с большей вероятностью вступают в контакт друг с другом и, следовательно, принимают участие в химических реакциях , поддерживающих жизнь. [32] С точки зрения своей молекулярной структуры вода представляет собой небольшую полярную молекулу с изогнутой формой, образованной полярными ковалентными связями двух атомов водорода (H) с одним атомом кислорода (O) (H 2 O). [32] Поскольку связи O–H являются полярными, атом кислорода имеет небольшой отрицательный заряд, а два атома водорода имеют небольшой положительный заряд. [32] Это полярное свойство воды позволяет ей притягивать другие молекулы воды через водородные связи, что делает воду связной . [32] Поверхностное натяжение возникает из-за силы сцепления, возникающей из-за притяжения между молекулами на поверхности жидкости. [32] Вода также является адгезивной , поскольку она способна прилипать к поверхности любых полярных или заряженных неводных молекул. [32] Вода плотнее в жидком состоянии , чем в твердом состоянии (или льда). [32] Это уникальное свойство воды позволяет льду плавать над жидкой водой, такой как пруды, озера и океаны, тем самым изолируя жидкость под ней от холодного воздуха над ней. [32] Вода обладает способностью поглощать энергию, что дает ей более высокую удельную теплоемкость , чем другие растворители, такие как этанол . [32] Таким образом, для разрыва водородных связей между молекулами воды и превращения жидкой воды в водяной пар требуется большое количество энергии . [32] Как молекула, вода не является полностью стабильной, поскольку каждая молекула воды непрерывно диссоциирует на ионы водорода и гидроксила , прежде чем снова превратиться в молекулу воды. [32] В чистой воде количество ионов водорода уравновешивает (или равно) количество гидроксильных ионов, в результате чего pH становится нейтральным.

Органические соединения — это молекулы, которые содержат углерод, связанный с другим элементом, таким как водород. [32] За исключением воды, почти все молекулы, составляющие каждый организм, содержат углерод. [32] [33] Углерод может образовывать ковалентные связи с четырьмя другими атомами, что позволяет ему образовывать разнообразные, большие и сложные молекулы. [32] [33] Например, один атом углерода может образовывать четыре одинарные ковалентные связи, такие как в метане , две двойные ковалентные связи , такие как в диоксиде углерода (CO 2 ), или тройную ковалентную связь , такую как в оксиде углерода (CO). Более того, углерод может образовывать очень длинные цепи взаимосвязанных углерод-углеродных связей, такие как октан , или кольцевые структуры, такие как глюкоза .
Простейшей формой органической молекулы является углеводород , который представляет собой большое семейство органических соединений, состоящих из атомов водорода, связанных с цепочкой атомов углерода. Углеводородный остов может быть заменен другими элементами, такими как кислород (O), водород (H), фосфор (P) и сера (S), которые могут изменить химическое поведение этого соединения. [32] Группы атомов, которые содержат эти элементы (O-, H-, P- и S-) и связаны с центральным атомом углерода или скелетом, называются функциональными группами . [32] Существует шесть основных функциональных групп, которые можно найти в организмах: аминогруппа , карбоксильная группа , карбонильная группа , гидроксильная группа , фосфатная группа и сульфгидрильная группа . [32]
В 1953 году эксперимент Миллера-Юри показал, что органические соединения могли синтезироваться абиотически в закрытой системе, имитирующей условия ранней Земли , тем самым предполагая, что сложные органические молекулы могли возникать спонтанно на ранней Земле (см. абиогенез ). [34] [32]

Макромолекулы — это большие молекулы, состоящие из более мелких субъединиц или мономеров . [35] Мономеры включают сахара, аминокислоты и нуклеотиды. [36] Углеводы включают мономеры и полимеры сахаров. [37] Липиды — это единственный класс макромолекул, которые не состоят из полимеров. Они включают стероиды , фосфолипиды и жиры, [36] в основном неполярные и гидрофобные (водоотталкивающие) вещества. [38] Белки — самые разнообразные из макромолекул. Они включают ферменты , транспортные белки , большие сигнальные молекулы, антитела и структурные белки . Основная единица (или мономер) белка — аминокислота . [35] В белках используется двадцать аминокислот. [35] Нуклеиновые кислоты — это полимеры нуклеотидов . [39] Их функция — хранить, передавать и выражать наследственную информацию. [36]
Клеточная теория утверждает, что клетки являются основными единицами жизни, что все живые существа состоят из одной или нескольких клеток, и что все клетки возникают из уже существующих клеток посредством деления клеток . [40] Большинство клеток очень малы, их диаметр составляет от 1 до 100 микрометров , и поэтому их можно увидеть только под световым или электронным микроскопом . [41] Обычно существует два типа клеток: эукариотические клетки, которые содержат ядро , и прокариотические клетки, которые его не содержат. Прокариоты — это одноклеточные организмы , такие как бактерии , тогда как эукариоты могут быть одноклеточными или многоклеточными. У многоклеточных организмов каждая клетка в теле организма в конечном итоге происходит из одной клетки в оплодотворенной яйцеклетке .

Каждая клетка заключена в клеточную мембрану , которая отделяет ее цитоплазму от внеклеточного пространства . [42] Клеточная мембрана состоит из липидного бислоя , включая холестерины , которые находятся между фосфолипидами, чтобы поддерживать их текучесть при различных температурах. Клеточные мембраны полупроницаемы , позволяя проходить через них небольшим молекулам, таким как кислород, углекислый газ и вода, ограничивая при этом движение более крупных молекул и заряженных частиц, таких как ионы . [43] Клеточные мембраны также содержат мембранные белки , включая интегральные мембранные белки , которые проходят через мембрану, выступая в качестве мембранных транспортеров , и периферические белки , которые свободно прикрепляются к внешней стороне клеточной мембраны, действуя как ферменты, формирующие клетку. [44] Клеточные мембраны участвуют в различных клеточных процессах, таких как клеточная адгезия , хранение электрической энергии и клеточная сигнализация , и служат поверхностью прикрепления для нескольких внеклеточных структур, таких как клеточная стенка , гликокаликс и цитоскелет .

В цитоплазме клетки находится множество биомолекул, таких как белки и нуклеиновые кислоты . [45] Помимо биомолекул, эукариотические клетки имеют специализированные структуры, называемые органеллами , которые имеют собственные липидные бислои или являются пространственными единицами. [46] Эти органеллы включают клеточное ядро , которое содержит большую часть ДНК клетки, или митохондрии , которые генерируют аденозинтрифосфат (АТФ) для питания клеточных процессов. Другие органеллы, такие как эндоплазматический ретикулум и аппарат Гольджи, играют роль в синтезе и упаковке белков соответственно. Биомолекулы, такие как белки, могут быть поглощены лизосомами , другой специализированной органеллой. Растительные клетки имеют дополнительные органеллы, которые отличают их от животных клеток, такие как клеточная стенка, которая обеспечивает поддержку растительной клетки, хлоропласты , которые собирают энергию солнечного света для производства сахара, и вакуоли , которые обеспечивают хранение и структурную поддержку, а также участвуют в воспроизводстве и расщеплении семян растений. [46] Эукариотические клетки также имеют цитоскелет, состоящий из микротрубочек , промежуточных филаментов и микрофиламентов , каждый из которых обеспечивает поддержку клетки и участвует в движении клетки и ее органелл. [46] С точки зрения их структурного состава, микротрубочки состоят из тубулина (например, α-тубулина и β-тубулина ), тогда как промежуточные филаменты состоят из фибриллярных белков. [46] Микрофиламенты состоят из молекул актина , которые взаимодействуют с другими цепями белков. [46]

Всем клеткам требуется энергия для поддержания клеточных процессов. Метаболизм — это набор химических реакций в организме. Три основные цели метаболизма: преобразование пищи в энергию для запуска клеточных процессов; преобразование пищи/топлива в мономерные строительные блоки; и устранение метаболических отходов . Эти катализируемые ферментами реакции позволяют организмам расти и размножаться, поддерживать свои структуры и реагировать на окружающую среду. Метаболические реакции можно разделить на катаболические — расщепление соединений (например, расщепление глюкозы до пирувата клеточным дыханием ); или анаболические — создание ( синтез ) соединений (таких как белки, углеводы, липиды и нуклеиновые кислоты). Обычно катаболизм высвобождает энергию, а анаболизм потребляет ее. Химические реакции метаболизма организованы в метаболические пути , в которых одно химическое вещество преобразуется через ряд шагов в другое химическое вещество, причем каждый шаг облегчается определенным ферментом. Ферменты имеют решающее значение для метаболизма, поскольку они позволяют организмам управлять желаемыми реакциями, требующими энергии, которая не произойдет сама по себе, связывая их со спонтанными реакциями , которые высвобождают энергию. Ферменты действуют как катализаторы — они позволяют реакции протекать быстрее, не потребляясь ею — уменьшая количество энергии активации, необходимой для превращения реагентов в продукты . Ферменты также позволяют регулировать скорость метаболической реакции, например, в ответ на изменения в среде клетки или на сигналы от других клеток.

Клеточное дыхание представляет собой набор метаболических реакций и процессов, которые происходят в клетках для преобразования химической энергии из питательных веществ в аденозинтрифосфат (АТФ) и последующего высвобождения отходов. [47] Реакции, участвующие в дыхании, являются катаболическими реакциями , которые расщепляют большие молекулы на более мелкие, высвобождая энергию. Дыхание является одним из ключевых способов, с помощью которых клетка высвобождает химическую энергию для подпитки клеточной активности. Общая реакция происходит в серии биохимических этапов, некоторые из которых являются окислительно-восстановительными реакциями. Хотя клеточное дыхание технически является реакцией горения , оно явно не похоже на нее, когда происходит в клетке, из-за медленного, контролируемого высвобождения энергии из серии реакций.
Сахар в форме глюкозы является основным питательным веществом, используемым клетками животных и растений для дыхания. Клеточное дыхание с участием кислорода называется аэробным дыханием, которое состоит из четырех стадий: гликолиза , цикла лимонной кислоты (или цикла Кребса), цепи переноса электронов и окислительного фосфорилирования . [48] Гликолиз — это метаболический процесс, который происходит в цитоплазме, в ходе которого глюкоза превращается в два пирувата , при этом одновременно производятся две чистые молекулы АТФ. [48] Затем каждый пируват окисляется до ацетил-КоА комплексом пируватдегидрогеназы , который также генерирует НАДН и углекислый газ. Ацетил-КоА входит в цикл лимонной кислоты, который происходит внутри митохондриальной матрицы. В конце цикла общий выход из 1 глюкозы (или 2 пируватов) составляет 6 НАДН, 2 ФАДН2 и 2 молекулы АТФ. Наконец, следующим этапом является окислительное фосфорилирование, которое у эукариот происходит в митохондриальных кристах . Окислительное фосфорилирование включает в себя цепь переноса электронов, которая представляет собой серию из четырех белковых комплексов , которые переносят электроны от одного комплекса к другому, тем самым высвобождая энергию из НАДН и ФАДН 2 , которая связана с перекачкой протонов (ионов водорода) через внутреннюю митохондриальную мембрану ( хемиосмос ), что создает протондвижущую силу . [48] Энергия от протондвижущей силы заставляет фермент АТФ-синтазу синтезировать больше АТФ путем фосфорилирования АДФ . Передача электронов заканчивается тем, что молекулярный кислород становится конечным акцептором электронов .
Если бы кислорода не было, пируват не метаболизировался бы клеточным дыханием, а подвергался бы процессу ферментации . Пируват не транспортируется в митохондрии, а остается в цитоплазме, где он преобразуется в отходы , которые могут быть удалены из клетки. Это служит цели окисления переносчиков электронов, чтобы они могли снова выполнять гликолиз, и удаления избытка пирувата. Ферментация окисляет NADH до NAD + , чтобы его можно было повторно использовать в гликолизе. При отсутствии кислорода ферментация предотвращает накопление NADH в цитоплазме и обеспечивает NAD + для гликолиза. Этот продукт отходов варьируется в зависимости от организма. В скелетных мышцах продуктом отходов является молочная кислота . Этот тип ферментации называется ферментацией молочной кислоты . При интенсивных упражнениях, когда энергетические потребности превышают энергопоставки, дыхательная цепь не может обработать все атомы водорода, присоединенные NADH. Во время анаэробного гликолиза NAD + регенерируется, когда пары водорода соединяются с пируватом, образуя лактат. Образование лактата катализируется лактатдегидрогеназой в обратимой реакции. Лактат также может использоваться в качестве непрямого предшественника гликогена печени. Во время восстановления, когда кислород становится доступным, NAD + присоединяется к водороду из лактата, образуя АТФ. У дрожжей отходами являются этанол и углекислый газ. Этот тип брожения известен как спиртовое или этаноловое брожение . АТФ, образующийся в этом процессе, производится путем фосфорилирования на уровне субстрата , которое не требует кислорода.

Фотосинтез — это процесс, используемый растениями и другими организмами для преобразования световой энергии в химическую энергию , которая впоследствии может быть высвобождена для питания метаболической активности организма посредством клеточного дыхания. Эта химическая энергия хранится в молекулах углеводов, таких как сахара, которые синтезируются из углекислого газа и воды. [49] [50] [51] В большинстве случаев кислород выделяется как побочный продукт. Большинство растений, водорослей и цианобактерий осуществляют фотосинтез, который в значительной степени отвечает за производство и поддержание содержания кислорода в атмосфере Земли и поставляет большую часть энергии, необходимой для жизни на Земле. [52]
Фотосинтез состоит из четырех стадий: поглощение света , перенос электронов, синтез АТФ и фиксация углерода . [48] Поглощение света является начальным этапом фотосинтеза, на котором энергия света поглощается пигментами хлорофилла , прикрепленными к белкам в мембранах тилакоидов . Поглощенная энергия света используется для удаления электронов от донора (воды) к первичному акцептору электронов, хинону , обозначенному как Q. На втором этапе электроны перемещаются от первичного акцептора электронов хинона через ряд переносчиков электронов, пока не достигнут конечного акцептора электронов, который обычно представляет собой окисленную форму НАДФ + , которая восстанавливается до НАДФН, процесс, который происходит в белковом комплексе, называемом фотосистемой I (PSI). Транспорт электронов сопряжен с движением протонов (или водорода) из стромы в мембрану тилакоида, что формирует градиент pH через мембрану, поскольку водород становится более концентрированным в просвете, чем в строме. Это аналогично протон-движущей силе, генерируемой через внутреннюю митохондриальную мембрану при аэробном дыхании. [48]
На третьем этапе фотосинтеза движение протонов по градиентам их концентрации из просвета тилакоида в строму через АТФ-синтазу сопряжено с синтезом АТФ той же АТФ-синтазой. [48] НАДФН и АТФ, генерируемые светозависимыми реакциями на втором и третьем этапах соответственно, обеспечивают энергию и электроны для запуска синтеза глюкозы путем фиксации атмосферного углекислого газа в существующих органических углеродных соединениях, таких как рибулозобисфосфат (РБФ) в последовательности светонезависимых (или темновых) реакций, называемых циклом Кальвина . [53]
Клеточная сигнализация (или коммуникация) — это способность клеток получать, обрабатывать и передавать сигналы с окружающей средой и с собой. [54] [55] Сигналы могут быть нехимическими, такими как свет, электрические импульсы и тепло, или химическими сигналами (или лигандами ), которые взаимодействуют с рецепторами , которые могут быть обнаружены встроенными в клеточную мембрану другой клетки или расположенными глубоко внутри клетки. [56] [55] Обычно существует четыре типа химических сигналов: аутокринные , паракринные , юкстакринные и гормоны . [56] При аутокринной сигнализации лиганд воздействует на ту же клетку, которая его высвобождает. Например, опухолевые клетки могут бесконтрольно размножаться, потому что они высвобождают сигналы, которые инициируют их собственное самоделение. При паракринной сигнализации лиганд диффундирует в близлежащие клетки и воздействует на них. Например, клетки мозга, называемые нейронами , высвобождают лиганды, называемые нейротрансмиттерами , которые диффундируют через синаптическую щель , чтобы связаться с рецептором на соседней клетке, такой как другой нейрон или мышечная клетка . При юкстакринной сигнализации существует прямой контакт между сигнальными и реагирующими клетками. Наконец, гормоны — это лиганды, которые перемещаются по кровеносной системе животных или сосудистой системе растений, чтобы достичь своих целевых клеток. Как только лиганд связывается с рецептором, он может влиять на поведение другой клетки, в зависимости от типа рецептора. Например, нейротрансмиттеры, которые связываются с инотропным рецептором, могут изменять возбудимость целевой клетки. Другие типы рецепторов включают рецепторы протеинкиназы (например, рецептор для гормона инсулина ) и рецепторы, сопряженные с G-белком . Активация рецепторов, сопряженных с G-белком, может инициировать каскады вторичных мессенджеров . Процесс, посредством которого химический или физический сигнал передается через клетку в виде серии молекулярных событий, называется трансдукцией сигнала .

Клеточный цикл представляет собой ряд событий, происходящих в клетке , которые заставляют ее делиться на две дочерние клетки. Эти события включают в себя дупликацию ее ДНК и некоторых ее органелл , а также последующее разделение ее цитоплазмы на две дочерние клетки в процессе, называемом делением клетки . [57] У эукариот (т. е. животных, растений, грибов и простейших клеток) существует два различных типа деления клеток: митоз и мейоз . [58] Митоз является частью клеточного цикла, в котором реплицированные хромосомы разделяются на два новых ядра. Деление клетки приводит к образованию генетически идентичных клеток, в которых сохраняется общее число хромосом. В целом, митозу (делению ядра) предшествует S-стадия интерфазы ( во время которой реплицируется ДНК), и за ним часто следуют телофаза и цитокинез ; который делит цитоплазму , органеллы и клеточную мембрану одной клетки на две новые клетки , содержащие примерно равные доли этих клеточных компонентов. Различные стадии митоза в совокупности определяют митотическую фазу цикла клеток животных — деление материнской клетки на две генетически идентичные дочерние клетки. [59] Клеточный цикл — это жизненно важный процесс, посредством которого одноклеточная оплодотворенная яйцеклетка развивается в зрелый организм, а также процесс, посредством которого обновляются волосы, кожа, клетки крови и некоторые внутренние органы . После деления клетки каждая из дочерних клеток начинает интерфазу нового цикла. В отличие от митоза, мейоз приводит к образованию четырех гаплоидных дочерних клеток, проходя один раунд репликации ДНК, за которым следуют два деления. [60] Гомологичные хромосомы разделяются в первом делении ( мейоз I ), а сестринские хроматиды разделяются во втором делении ( мейоз II ). Оба этих цикла деления клеток используются в процессе полового размножения в какой-то момент их жизненного цикла. Считается, что оба присутствуют у последнего общего предка эукариот.
Прокариоты (т. е. археи и бактерии) также могут подвергаться делению клеток (или бинарному делению ). В отличие от процессов митоза и мейоза у эукариот, бинарное деление у прокариот происходит без образования веретенного аппарата на клетке. Перед бинарным делением ДНК в бактерии плотно свернута. После того, как она раскручивается и дуплицируется, она тянется к отдельным полюсам бактерии, поскольку она увеличивается в размере, чтобы подготовиться к разделению. Рост новой клеточной стенки начинает разделять бактерию (вызванный полимеризацией FtsZ и образованием «Z-кольца»). [61] Новая клеточная стенка ( септум ) полностью развивается, что приводит к полному разделению бактерии. Новые дочерние клетки имеют плотно скрученные стержни ДНК, рибосомы и плазмиды .
Мейоз является центральной особенностью полового размножения у эукариот, и наиболее фундаментальной функцией мейоза , по-видимому, является сохранение целостности генома , который передается потомству родителями. [62] [63] Два аспекта полового размножения , мейотическая рекомбинация и ауткроссинг , вероятно, поддерживаются соответственно адаптивными преимуществами рекомбинационной репарации повреждений геномной ДНК и генетической комплементации , которая маскирует экспрессию вредных рецессивных мутаций . [64]
Благоприятный эффект генетической комплементарности, полученный в результате ауткроссинга (перекрестного оплодотворения), также называется гибридной силой или гетерозисом. Чарльз Дарвин в своей книге 1878 года « Влияние перекрестного и самооплодотворения в растительном мире» [65] в начале главы XII отметил: «Первый и самый важный из выводов, который можно сделать из наблюдений, приведенных в этом томе, заключается в том, что обычно перекрестное оплодотворение полезно, а самооплодотворение часто вредно, по крайней мере, с растениями, на которых я экспериментировал». Генетическая изменчивость , часто возникающая как побочный продукт полового размножения, может обеспечить долгосрочные преимущества тем половым линиям, которые участвуют в ауткроссинге . [64]

Генетика — это научное изучение наследования. [66] [67] [68] Менделевское наследование , в частности, — это процесс, посредством которого гены и признаки передаются от родителей к потомству. [28] Оно имеет несколько принципов. Первый заключается в том, что генетические характеристики, аллели , являются дискретными и имеют альтернативные формы (например, фиолетовый против белого или высокий против карликового), каждый из которых наследуется от одного из двух родителей. Основанный на законе доминирования и единообразия , который гласит, что некоторые аллели являются доминантными , а другие рецессивными ; организм, имеющий по крайней мере один доминантный аллель, будет демонстрировать фенотип этого доминантного аллеля. Во время формирования гамет аллели для каждого гена разделяются, так что каждая гамета несет только один аллель для каждого гена. Гетерозиготные особи производят гаметы с равной частотой двух аллелей. Наконец, закон независимого распределения гласит, что гены разных признаков могут разделяться независимо во время формирования гамет, т. е. гены не связаны. Исключением из этого правила могут быть признаки, сцепленные с полом . Тестовые скрещивания могут быть выполнены для экспериментального определения основного генотипа организма с доминирующим фенотипом. [69] Решетка Паннета может быть использована для прогнозирования результатов тестового скрещивания. Хромосомная теория наследования , которая утверждает, что гены находятся на хромосомах, была поддержана экспериментами Томаса Моргана с плодовыми мушками , которые установили половую связь между цветом глаз и полом у этих насекомых. [70]

Ген — это единица наследственности , которая соответствует участку дезоксирибонуклеиновой кислоты (ДНК), которая несет генетическую информацию, которая контролирует форму или функцию организма. ДНК состоит из двух полинуклеотидных цепей, которые обвиваются друг вокруг друга, образуя двойную спираль . [71] Она встречается в виде линейных хромосом у эукариот и кольцевых хромосом у прокариот . Набор хромосом в клетке в совокупности известен как ее геном . У эукариот ДНК в основном находится в ядре клетки . [72] У прокариот ДНК содержится в нуклеоиде . [73] Генетическая информация содержится в генах, а полная сборка в организме называется его генотипом . [74] Репликация ДНК — это полуконсервативный процесс, при котором каждая нить служит шаблоном для новой нити ДНК. [71] Мутации — это наследуемые изменения в ДНК. [71] Они могут возникать спонтанно в результате ошибок репликации, которые не были исправлены корректурой, или могут быть вызваны мутагеном окружающей среды , таким как химикат (например, азотистая кислота , бензопирен ) или радиация (например, рентгеновские лучи , гамма-лучи , ультрафиолетовое излучение , частицы, испускаемые нестабильными изотопами). [71] Мутации могут приводить к фенотипическим эффектам, таким как потеря функции, приобретение функции и условные мутации. [71] Некоторые мутации полезны, так как они являются источником генетической изменчивости для эволюции. [71] Другие вредны, если они приводят к потере функции генов, необходимых для выживания. [71]

Экспрессия генов — это молекулярный процесс, посредством которого генотип , закодированный в ДНК, приводит к наблюдаемому фенотипу в белках тела организма. Этот процесс обобщен центральной догмой молекулярной биологии , сформулированной Фрэнсисом Криком в 1958 году. [75] [76] [77] Согласно Центральной догме, генетическая информация передается от ДНК к РНК и белку. Существует два процесса экспрессии генов: транскрипция (ДНК в РНК) и трансляция (РНК в белок). [78]
Регулирование экспрессии генов факторами окружающей среды и на разных стадиях развития может происходить на каждом этапе процесса, например, транскрипции , сплайсинга РНК , трансляции и посттрансляционной модификации белка. [79] На экспрессию генов может влиять положительная или отрицательная регуляция, в зависимости от того, какой из двух типов регуляторных белков, называемых факторами транскрипции, связывается с последовательностью ДНК вблизи или на промоторе. [79] Кластер генов, которые разделяют один и тот же промотор, называется опероном , он встречается в основном у прокариот и некоторых низших эукариот (например, Caenorhabditis elegans ). [79] [80] При положительной регуляции экспрессии генов активатором является фактор транскрипции, который стимулирует транскрипцию, когда он связывается с последовательностью вблизи или на промоторе. Отрицательная регуляция происходит, когда другой фактор транскрипции, называемый репрессором, связывается с последовательностью ДНК, называемой оператором , которая является частью оперона, чтобы предотвратить транскрипцию. Репрессоры могут быть ингибированы соединениями, называемыми индукторами (например, аллолактозой ), тем самым позволяя транскрипции происходить. [79] Конкретные гены, которые могут быть активированы индукторами, называются индуцируемыми генами , в отличие от конститутивных генов , которые почти постоянно активны. [79] В отличие от обоих, структурные гены кодируют белки, которые не участвуют в регуляции генов. [79] В дополнение к регуляторным событиям, включающим промотор, экспрессия генов также может регулироваться эпигенетическими изменениями хроматина , который представляет собой комплекс ДНК и белка, обнаруженный в эукариотических клетках. [79]
Развитие — это процесс, посредством которого многоклеточный организм (растение или животное) проходит через ряд изменений, начиная с одной клетки и принимая различные формы, характерные для его жизненного цикла. [81] В основе развития лежат четыре ключевых процесса: определение , дифференциация , морфогенез и рост. Определение определяет судьбу развития клетки, которая становится более ограниченной в процессе развития. Дифференциация — это процесс, посредством которого специализированные клетки возникают из менее специализированных клеток, таких как стволовые клетки . [82] [83] Стволовые клетки — это недифференцированные или частично дифференцированные клетки , которые могут дифференцироваться в различные типы клеток и размножаться неограниченно, чтобы производить больше тех же стволовых клеток. [84] Клеточная дифференциация резко изменяет размер, форму, мембранный потенциал , метаболическую активность и восприимчивость клетки к сигналам, что в значительной степени обусловлено строго контролируемыми модификациями в экспрессии генов и эпигенетике . За некоторыми исключениями, клеточная дифференциация почти никогда не включает изменение самой последовательности ДНК . [85] Таким образом, разные клетки могут иметь очень разные физические характеристики, несмотря на то, что имеют один и тот же геном . Морфогенез, или развитие формы тела, является результатом пространственных различий в экспрессии генов. [81] Небольшая часть генов в геноме организма, называемая набором генетического инструментария развития, контролирует развитие этого организма. Эти гены набора инструментов высококонсервативны среди типов , что означает, что они являются древними и очень похожими в широко разделенных группах животных. Различия в развертывании генов набора инструментов влияют на план тела и количество, идентичность и рисунок частей тела. Среди наиболее важных генов набора инструментов находятся гены Hox . Гены Hox определяют, где повторяющиеся части, такие как многочисленные позвонки змей, будут расти в развивающемся эмбрионе или личинке. [86]

Эволюция является центральным организующим понятием в биологии. Это изменение наследуемых характеристик популяций в последовательных поколениях . [87] [88] При искусственном отборе животные селективно разводились по определенным признакам. [89] Учитывая, что признаки наследуются, популяции содержат разнообразный набор признаков, а воспроизводство способно увеличить любую популяцию, Дарвин утверждал, что в естественном мире именно природа играла роль людей в отборе определенных признаков. [89] Дарвин сделал вывод, что особи, обладающие наследуемыми признаками, лучше приспособленными к своей среде, с большей вероятностью выживут и произведут больше потомства, чем другие особи. [89] Он также сделал вывод, что это приведет к накоплению благоприятных признаков в последовательных поколениях, тем самым увеличивая соответствие между организмами и их средой. [90] [91] [92] [89] [93]
Вид — это группа организмов, которые спариваются друг с другом, а видообразование — это процесс, посредством которого одна линия разделяется на две линии в результате независимой эволюции друг от друга. [94] Для того чтобы произошло видообразование, должна быть репродуктивная изоляция . [94] Репродуктивная изоляция может быть результатом несовместимости между генами, как описано в модели Бейтсона-Добжанского-Мюллера . Репродуктивная изоляция также имеет тенденцию увеличиваться с генетической дивергенцией . Видообразование может происходить, когда существуют физические барьеры, разделяющие предковый вид, процесс, известный как аллопатрическое видообразование . [94]

Филогенетика — это эволюционная история определенной группы организмов или их генов. [95] Ее можно представить с помощью филогенетического дерева , диаграммы, показывающей линии происхождения организмов или их генов. Каждая линия, нарисованная на оси времени дерева, представляет собой линию потомков определенного вида или популяции. Когда линия разделяется на две, она представляется в виде вилки или раскола на филогенетическом дереве. [95] Филогенетические деревья являются основой для сравнения и группировки различных видов. [95] Различные виды, которые имеют общую черту, унаследованную от общего предка, описываются как имеющие гомологичные черты (или синапоморфию ). [96] [97] [95] Филогенетика обеспечивает основу биологической классификации. [95] Эта система классификации основана на рангах, причем наивысший ранг — это домен, за которым следуют царство , тип , класс , порядок , семейство , род и вид . [95] Все организмы можно классифицировать как принадлежащие к одному из трех доменов : археи (первоначально архебактерии), бактерии (первоначально эубактерии) или эукариоты (включает грибы, растения и животные царства). [98]
История жизни на Земле прослеживает, как организмы эволюционировали от самого раннего появления жизни до наших дней. Земля образовалась около 4,5 миллиардов лет назад, и вся жизнь на Земле, как живая, так и вымершая, произошла от последнего универсального общего предка , который жил около 3,5 миллиардов лет назад . [99] [100] Геологи разработали геологическую шкалу времени , которая делит историю Земли на основные периоды, начиная с четырех эонов ( гадей , архей , протерозой и фанерозой ), первые три из которых в совокупности известны как докембрий , который длился приблизительно 4 миллиарда лет. [101] Каждый эон можно разделить на эры, при этом фанерозойский эон, начавшийся 539 миллионов лет назад [102] , подразделяется на палеозойскую , мезозойскую и кайнозойскую эры. [101] Эти три эры вместе составляют одиннадцать периодов ( кембрийский , ордовикский , силурийский , девонский , каменноугольный , пермский , триасовый , юрский , меловой , третичный и четвертичный ). [101]
Сходства среди всех известных современных видов указывают на то, что они разошлись в процессе эволюции от своего общего предка. [103] Биологи рассматривают повсеместность генетического кода как доказательство всеобщего общего происхождения всех бактерий , архей и эукариот . [104] [10] [105] [106] Микробные маты сосуществующих бактерий и архей были доминирующей формой жизни в раннем архейском эоне, и многие из основных шагов ранней эволюции, как полагают, имели место в этой среде. [107] Самые ранние свидетельства существования эукариот датируются 1,85 миллиарда лет назад, [108] [109] и хотя они могли присутствовать и раньше, их диверсификация ускорилась, когда они начали использовать кислород в своем метаболизме . Позже, около 1,7 миллиарда лет назад, начали появляться многоклеточные организмы с дифференцированными клетками , выполняющими специализированные функции. [110]
Водорослеподобные многоклеточные наземные растения датируются примерно 1 миллиардом лет назад, [111] хотя данные свидетельствуют о том, что микроорганизмы сформировали самые ранние наземные экосистемы , по крайней мере 2,7 миллиарда лет назад. [112] Считается, что микроорганизмы проложили путь к появлению наземных растений в ордовикский период. Наземные растения были настолько успешны, что, как полагают, внесли свой вклад в позднедевонское вымирание . [113]
Биота Эдиакары появилась в эдиакарский период, [114] в то время как позвоночные , наряду с большинством других современных типов , возникли около 525 миллионов лет назад во время кембрийского взрыва . [115] В пермский период синапсиды , включая предков млекопитающих , доминировали на суше, [116] но большая часть этой группы вымерла во время пермско-триасового вымирания 252 миллиона лет назад. [117] Во время восстановления после этой катастрофы архозавры стали самыми многочисленными наземными позвоночными; [118] одна группа архозавров, динозавры, доминировала в юрский и меловой периоды. [119] После того, как мел-палеогеновое вымирание 66 миллионов лет назад уничтожило нептичьих динозавров, [120] млекопитающие быстро увеличились в размерах и разнообразии . [121] Такие массовые вымирания могли ускорить эволюцию, предоставив возможности для диверсификации новых групп организмов. [122]

Бактерии — это тип клеток , которые составляют большую область прокариотических микроорганизмов . Обычно длиной в несколько микрометров , бактерии имеют ряд форм , от сфер до палочек и спиралей . Бактерии были одними из первых форм жизни, появившихся на Земле, и присутствуют в большинстве ее местообитаний . Бактерии населяют почву, воду, кислые горячие источники , радиоактивные отходы [ 123] и глубокую биосферу земной коры . Бактерии также живут в симбиотических и паразитических отношениях с растениями и животными. Большинство бактерий не были охарактеризованы, и только около 27 процентов бактериальных типов имеют виды, которые можно выращивать в лаборатории. [124]

Археи составляют другой домен прокариотических клеток и изначально были классифицированы как бактерии, получив название архебактерии (в царстве архебактерий ), термин, который вышел из употребления. [125] Архейные клетки обладают уникальными свойствами, отделяющими их от двух других доменов , бактерий и эукариот . Археи далее делятся на несколько признанных типов . Археи и бактерии, как правило, похожи по размеру и форме, хотя некоторые археи имеют очень разные формы, такие как плоские и квадратные клетки Haloquadratum walsbyi . [126] Несмотря на это морфологическое сходство с бактериями, археи обладают генами и несколькими метаболическими путями , которые более тесно связаны с таковыми у эукариот, особенно для ферментов , участвующих в транскрипции и трансляции . Другие аспекты биохимии архей уникальны, такие как их зависимость от эфирных липидов в их клеточных мембранах , [127] включая археолы . Археи используют больше источников энергии, чем эукариоты: они варьируются от органических соединений , таких как сахара, до аммиака , ионов металлов или даже водорода . Солеустойчивые археи ( Haloarchaea ) используют солнечный свет в качестве источника энергии, а другие виды архей фиксируют углерод , но в отличие от растений и цианобактерий , ни один известный вид архей не делает и то, и другое. Археи размножаются бесполым путем путем бинарного деления , фрагментации или почкования ; в отличие от бактерий, ни один известный вид архей не образует эндоспоры .
Первые обнаруженные археи были экстремофилами , живущими в экстремальных условиях, таких как горячие источники и соленые озера без других организмов. Улучшенные молекулярные инструменты обнаружения привели к обнаружению архей практически в каждой среде обитания , включая почву, океаны и болота . Археи особенно многочисленны в океанах, а археи в планктоне могут быть одной из самых распространенных групп организмов на планете.
Археи играют важную роль в жизни Земли . Они являются частью микробиоты всех организмов. В микробиоме человека они играют важную роль в кишечнике , во рту и на коже. [128] Их морфологическое, метаболическое и географическое разнообразие позволяет им играть множество экологических ролей: фиксация углерода; круговорот азота; оборот органических соединений; и поддержание микробных симбиотических и синтрофных сообществ, например. [129]

Предполагается, что эукариоты отделились от архей, за чем последовал их эндосимбиоз с бактериями (или симбиогенез ), который дал начало митохондриям и хлоропластам, которые теперь являются частью современных эукариотических клеток. [130] Основные линии эукариот диверсифицировались в докембрии около 1,5 миллиарда лет назад и могут быть разделены на восемь основных клад : альвеоляты , экскаваты , страменопилы , растения, ризарии , амебозои , грибы и животные. [130] Пять из этих клад известны под общим названием протисты , которые в основном представляют собой микроскопические эукариотические организмы, не являющиеся растениями, грибами или животными. [130] Хотя вполне вероятно, что протисты имеют общего предка ( последнего общего предка эукариот ), [131] протисты сами по себе не составляют отдельную кладу, поскольку некоторые протисты могут быть более тесно связаны с растениями, грибами или животными, чем с другими протистами. Подобно таким группам, как водоросли , беспозвоночные или простейшие , группа протистов не является формальной таксономической группой, но используется для удобства. [130] [132] Большинство протистов одноклеточные; их называют микробными эукариотами. [130]
Растения в основном являются многоклеточными организмами , преимущественно фотосинтезирующими эукариотами царства Plantae , которое исключает грибы и некоторые водоросли . Растительные клетки произошли от эндосимбиоза цианобактерий в ранних эукариот около миллиарда лет назад, что дало начало хлоропластам. [133] Первые несколько клад, возникших после первичного эндосимбиоза, были водными, и большинство водных фотосинтетических эукариотических организмов в совокупности описываются как водоросли, что является термином для удобства, поскольку не все водоросли тесно связаны. [133] Водоросли включают в себя несколько отдельных клад, таких как глаукофиты , которые представляют собой микроскопические пресноводные водоросли, которые могли напоминать по форме раннего одноклеточного предка Plantae. [133] В отличие от глаукофитов, другие клады водорослей, такие как красные и зеленые водоросли, являются многоклеточными. Зелёные водоросли включают три основных клада: хлорофиты , колеохэтофиты и каменистые водоросли . [133]
Грибы — это эукариоты, которые переваривают пищу вне своего тела, [134] выделяя пищеварительные ферменты, которые расщепляют большие молекулы пищи перед тем, как впитать их через клеточные мембраны. Многие грибы также являются сапробионтами , питаясь мертвым органическим веществом, что делает их важными редуцентами в экологических системах. [134]
Животные — многоклеточные эукариоты. За редкими исключениями, животные потребляют органический материал , дышат кислородом , способны двигаться , могут размножаться половым путем и расти из полой сферы клеток , бластулы , во время эмбрионального развития . Описано более 1,5 миллионов ныне живущих видов животных , из которых около 1 миллиона — насекомые , но, по оценкам, всего существует более 7 миллионов видов животных. Они имеют сложные взаимодействия друг с другом и своей средой, образуя запутанные пищевые сети . [135]

Вирусы — это субмикроскопические инфекционные агенты , которые размножаются внутри клеток организмов . [136] Вирусы заражают все типы форм жизни , от животных и растений до микроорганизмов , включая бактерии и археи . [137] [138] Более 6000 видов вирусов были подробно описаны. [139] Вирусы встречаются почти в каждой экосистеме на Земле и являются наиболее многочисленным типом биологической сущности. [140] [141]
Происхождение вирусов в эволюционной истории жизни неясно: некоторые из них могли произойти от плазмид — фрагментов ДНК, которые могут перемещаться между клетками, — в то время как другие могли произойти от бактерий. В эволюции вирусы являются важным средством горизонтального переноса генов , что увеличивает генетическое разнообразие способом, аналогичным половому размножению . [142] Поскольку вирусы обладают некоторыми, но не всеми характеристиками жизни, их описывают как «организмы на грани жизни», [143] и как саморепликаторы . [144]
Экология – это изучение распространения и распространенности жизни, взаимодействия организмов с окружающей средой . [145]
Сообщество живых ( биотических ) организмов в сочетании с неживыми ( абиотическими ) компонентами (например, вода , свет, радиация, температура, влажность , атмосфера , кислотность и почва) их среды обитания называется экосистемой . [ 146] [147] [148] Эти биотические и абиотические компоненты связаны между собой посредством питательных циклов и потоков энергии. [149] Энергия от солнца поступает в систему через фотосинтез и включается в растительную ткань. Питаясь растениями и друг другом, животные перемещают вещество и энергию через систему. Они также влияют на количество присутствующей растительной и микробной биомассы . Разлагая мертвое органическое вещество , редуценты высвобождают углерод обратно в атмосферу и облегчают круговорот питательных веществ , преобразуя питательные вещества, хранящиеся в мертвой биомассе, обратно в форму, которая может быть легко использована растениями и другими микробами. [150]

Популяция — это группа организмов одного вида , которая занимает территорию и размножается из поколения в поколение. [151] [152] [153] [154] [155] Размер популяции можно оценить, умножив плотность популяции на площадь или объем. Грузоподъемность среды — это максимальный размер популяции вида, который может поддерживаться этой конкретной средой, учитывая пищу, среду обитания , воду и другие доступные ресурсы . [156] Грузоподъемность популяции может зависеть от изменяющихся условий окружающей среды, таких как изменения в доступности ресурсов и стоимости их содержания. В человеческих популяциях новые технологии , такие как Зеленая революция, помогли со временем увеличить грузоподъемность Земли для людей, что загнало в тупик попытки предсказания надвигающегося сокращения численности населения, самым известным из которых было предсказано Томасом Мальтусом в 18 веке. [151]

Сообщество — это группа популяций видов, занимающих одну и ту же географическую область в одно и то же время. [158] Биологическое взаимодействие — это эффект, который оказывают друг на друга пара организмов , живущих вместе в сообществе. Они могут быть как одного вида (внутривидовые взаимодействия), так и разных видов (межвидовые взаимодействия). Эти эффекты могут быть краткосрочными, как опыление и хищничество , или долгосрочными; оба часто сильно влияют на эволюцию вовлеченных видов. Долгосрочное взаимодействие называется симбиозом . Симбиозы варьируются от мутуализма , выгодного для обоих партнеров, до конкуренции , вредной для обоих партнеров. [159] Каждый вид участвует в качестве потребителя, ресурса или и того, и другого во взаимодействиях потребитель–ресурс , которые образуют ядро пищевых цепей или пищевых сетей . [160] В любой пищевой сети существуют различные трофические уровни , самый низкий уровень — это первичные производители (или автотрофы ), такие как растения и водоросли, которые преобразуют энергию и неорганические материалы в органические соединения , которые затем могут использоваться остальной частью сообщества. [52] [161] [162] На следующем уровне находятся гетеротрофы , которые являются видами, которые получают энергию путем расщепления органических соединений из других организмов. [160] Гетеротрофы, которые потребляют растения, являются первичными потребителями (или травоядными ), тогда как гетеротрофы, которые потребляют травоядных, являются вторичными потребителями (или плотоядными ). А те, которые едят вторичных потребителей, являются третичными потребителями и так далее. Всеядные гетеротрофы способны потреблять на нескольких уровнях. Наконец, есть редуценты , которые питаются отходами или мертвыми телами организмов. [160] В среднем, общее количество энергии, включенной в биомассу трофического уровня за единицу времени, составляет около одной десятой энергии трофического уровня, которую он потребляет. Отходы и мертвый материал, используемые редуцентами, а также тепло, потерянное в результате метаболизма, составляют остальные девяносто процентов энергии, которая не потребляется следующим трофическим уровнем. [163]
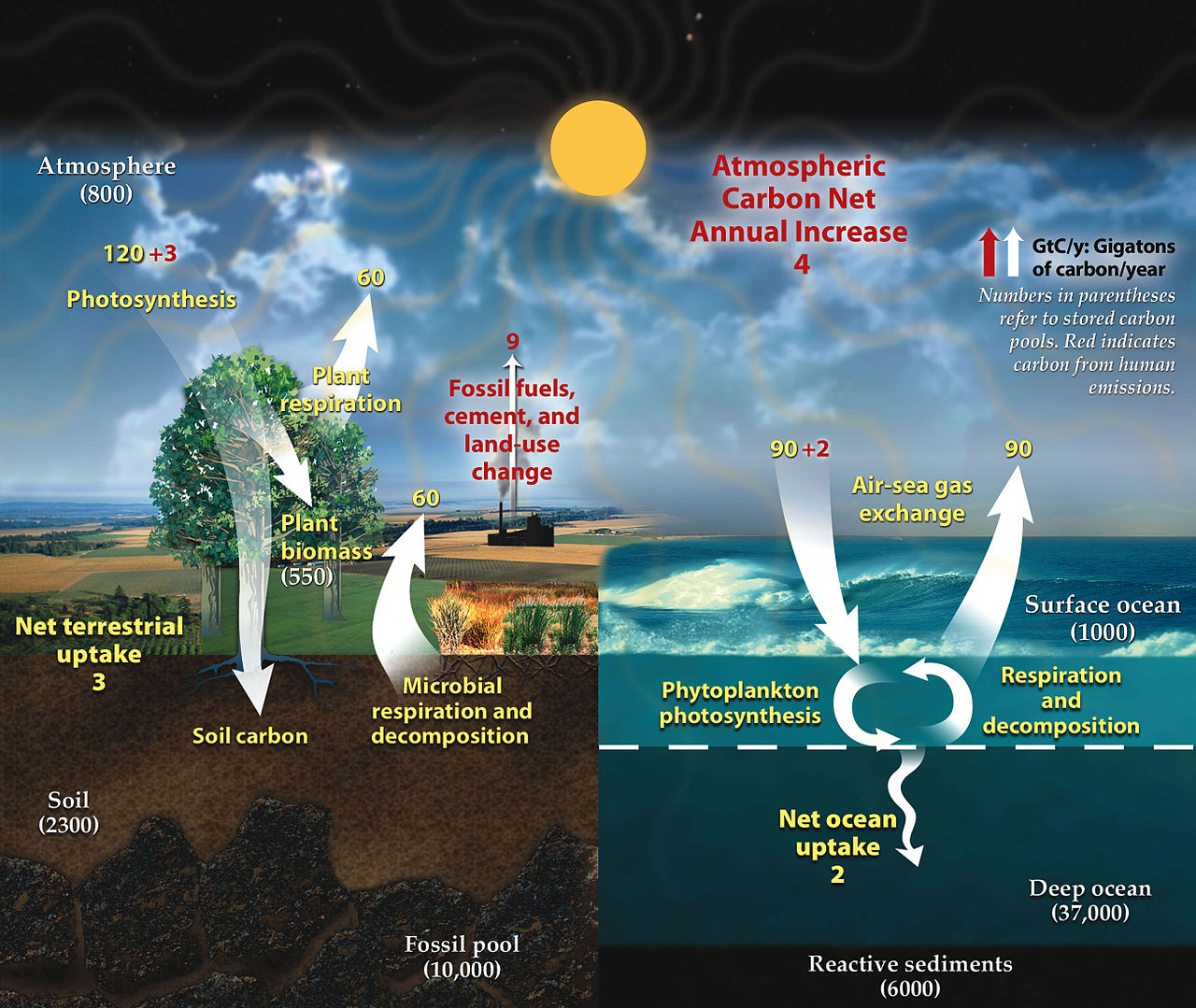
В глобальной экосистеме или биосфере материя существует в виде различных взаимодействующих отсеков, которые могут быть биотическими или абиотическими, а также доступными или недоступными, в зависимости от их форм и местонахождения. [165] Например, материя из наземных автотрофов является как биотической, так и доступной для других организмов, тогда как материя в горных породах и минералах является абиотической и недоступной. Биогеохимический цикл — это путь, по которому определенные элементы материи переворачиваются или перемещаются через биотические ( биосферу ) и абиотические ( литосферу , атмосферу и гидросферу ) отсеки Земли. Существуют биогеохимические циклы для азота , углерода и воды .
Биология сохранения — это изучение сохранения биоразнообразия Земли с целью защиты видов , их среды обитания и экосистем от чрезмерных темпов вымирания и эрозии биотических взаимодействий. [166] [167] [168] Она занимается факторами, которые влияют на поддержание, потерю и восстановление биоразнообразия, а также наукой поддержания эволюционных процессов, которые порождают генетическое , популяционное, видовое и экосистемное разнообразие. [169] [170] [171] [172] Беспокойство возникает из-за оценок, предполагающих, что до 50% всех видов на планете исчезнут в течение следующих 50 лет, [173] что способствовало нищете, голоду и сбросит ход эволюции на этой планете. [174] [175] Биоразнообразие влияет на функционирование экосистем, которые предоставляют различные услуги , от которых зависят люди. Биологи по охране природы исследуют и обучают тенденциям потери биоразнообразия , вымиранию видов и негативному влиянию, которое они оказывают на наши возможности поддерживать благосостояние человеческого общества. Организации и граждане реагируют на текущий кризис биоразнообразия с помощью планов действий по охране природы, которые направляют исследовательские, мониторинговые и образовательные программы, которые затрагивают проблемы на местном и глобальном уровнях. [176] [169] [170] [171]
{{cite web}}: CS1 maint: unfit URL (link)Это первоначальное включение углерода в органические соединения известно как фиксация углерода.
могли с уверенностью сказать, что формы изменяются, и что естественный отбор является важной силой для изменений. Однако они ничего не могли сказать о том, как это изменение осуществляется. Как изменяются тела или части тела, или как возникают новые структуры, оставалось полной загадкой.
сообщество животных, растений или людей, среди членов которого происходит скрещивание
– синтез организмами органических химических соединений, особенно углеводов, из углекислого газа с использованием энергии, получаемой от света, а не путем окисления химических соединений.
Ссылки на журналы
{kind=link}
{kind=link}