
Ботаника , также называемая наукой о растениях (или науками о растениях ), биологией растений или фитологией , — это наука о жизни растений и раздел биологии . Ботаник , ученый-растениевод или фитолог — это ученый , специализирующийся в этой области. Термин «ботаника» происходит от древнегреческого слова βοτάνη ( botanē ), означающего « пастбище », « травы », « трава » или « корм »; βοτάνη , в свою очередь, происходит от βόσκειν ( боскейн ), «кормить» или « пастись ». [1] [2] [3] Традиционно ботаника также включала изучение грибов и водорослей микологов и психологов соответственно, при этом изучение этих трех групп организмов остается в сфере интересов Международного ботанического конгресса . В настоящее время ботаники (в строгом смысле этого слова) изучают около 410 000 видов наземных растений , из которых около 391 000 видов — сосудистые растения (в том числе около 369 000 видов цветковых растений ) [4] и около 20 000 — мохообразные . [5]
Ботаника зародилась в доисторические времена как травничество , когда древние люди пытались идентифицировать – а затем выращивать – растения, которые были съедобными, ядовитыми и, возможно, лекарственными, что сделало ее одним из первых начинаний человечества. Средневековые лечебные сады , часто пристроенные к монастырям , содержали растения, возможно, имеющие лечебную ценность. Они были предшественниками первых ботанических садов при университетах , основанных с 1540-х годов. Одним из первых был Падуанский ботанический сад . Эти сады способствовали академическому изучению растений. Усилия по каталогизации и описанию их коллекций положили начало систематике растений и привели в 1753 году к биномиальной системе номенклатуры Карла Линнея , которая используется и по сей день для обозначения всех биологических видов.
В 19 и 20 веках были разработаны новые методы изучения растений, включая методы оптической микроскопии и визуализации живых клеток , электронную микроскопию , анализ числа хромосом , химии растений , а также структуры и функций ферментов и других белков . В последние два десятилетия 20-го века ботаники использовали методы молекулярно-генетического анализа , включая геномику и протеомику , а также последовательности ДНК , чтобы более точно классифицировать растения.
Современная ботаника — это широкий междисциплинарный предмет, в который внесены вклады и идеи большинства других областей науки и техники. Темы исследований включают изучение структуры растений , роста и дифференциации, размножения , биохимии и первичного метаболизма , химических продуктов, развития , болезней , эволюционных взаимоотношений , систематики и таксономии растений . Доминирующими темами в науке о растениях XXI века являются молекулярная генетика и эпигенетика , изучающие механизмы и контроль экспрессии генов во время дифференцировки растительных клеток и тканей . Ботанические исследования имеют разнообразные применения в обеспечении основных продуктов питания , таких материалов, как древесина , нефть , каучук, волокно и лекарства, в современном садоводстве , сельском и лесном хозяйстве , размножении растений , селекции и генетической модификации , в синтезе химикатов и сырья для строительства и производство энергии, управление окружающей средой и поддержание биоразнообразия .

Ботаника возникла как травничество , изучение и использование растений из-за их возможных лечебных свойств . [6] Ранняя история ботаники включает в себя множество древних сочинений и классификаций растений. Примеры ранних ботанических работ были найдены в древних текстах из Индии, датируемых ранее 1100 г. до н. э., [7] [8] Древнего Египта , [9] в архаичных древнеиранских авестийских сочинениях и в произведениях из Китая предположительно до 221 г. до н. э. [7] [10]
Современная ботаника уходит корнями в Древнюю Грецию , а именно к Теофрасту ( ок. 371–287 до н. э.), ученику Аристотеля , который изобрел и описал многие из ее принципов и широко известен в научном сообществе как «отец ботаники». [11] Его основные работы, «Исследование растений » и «О причинах возникновения растений» , представляют собой наиболее важный вклад в ботаническую науку до средневековья , почти семнадцать веков спустя. [11] [12]
Еще одна работа из Древней Греции, оказавшая раннее влияние на ботанику, — это De materia medica , пятитомная энциклопедия о предварительной фитотерапии, написанная в середине первого века греческим врачом и фармакологом Педаниусом Диоскоридом . De materia medica широко читалась на протяжении более 1500 лет. [13] Важные вклады средневекового мусульманского мира включают « Набатейское сельское хозяйство » Ибн Вахшии , «Книгу растений » Абу Ханифы Динавари (828–896) и «Классификацию почв» Ибн Бассала . В начале 13 века Абу аль-Аббас ан-Набати и Ибн аль-Байтар (ум. 1248) писали по ботанике систематическим и научным образом. [14] [15] [16]
В середине 16 века при ряде итальянских университетов были основаны ботанические сады . Ботанический сад Падуи 1545 года обычно считается первым, который до сих пор находится на своем первоначальном месте. Эти сады продолжили практическую ценность более ранних «физических садов», часто связанных с монастырями, в которых растения выращивались для предполагаемых медицинских целей. Они поддержали развитие ботаники как академического предмета. Были прочитаны лекции о растениях, выращиваемых в садах. Ботанические сады появились в Северной Европе гораздо позже; первым в Англии был Ботанический сад Оксфордского университета в 1621 году. [17]
Немецкий врач Леонхарт Фукс (1501–1566) был одним из «трех немецких отцов ботаники» наряду с теологом Отто Брунфельсом (1489–1534) и врачом Иеронимом Боком (1498–1554) (также называемым Иеронимом Козелком). [18] [19] Фукс и Брунфельс отошли от традиции копирования более ранних работ, чтобы сделать собственные оригинальные наблюдения. Бок создал собственную систему классификации растений.
Врач Валериус Корд (1515–1544) написал ботанически и фармакологически важную книгу « Historia Plantarum» в 1544 году и фармакопею непреходящего значения « Dispensatorium » в 1546 году . [20] Натуралист Конрад фон Геснер (1516–1565) и травник Джон Герард (1545–1545–1545 гг.) около 1611 г. ) опубликовал травники, освещающие предполагаемое медицинское использование растений. Натуралист Улиссе Альдрованди (1522–1605) считался отцом естествознания , включавшего изучение растений. В 1665 году, используя первый микроскоп, эрудит Роберт Гук обнаружил клетки (термин, который он придумал) в пробке , а некоторое время спустя и в живых растительных тканях. [21]

В XVIII веке были разработаны системы идентификации растений , сравнимые с дихотомическими ключами , в которых неопознанные растения помещаются в таксономические группы (например, семейство, род и вид) путем серии выборов между парами признаков . Выбор и последовательность символов могут быть искусственными в ключах, предназначенных исключительно для идентификации ( диагностические ключи ), или более тесно связанными с естественным или филетическим порядком таксонов в синоптических ключах. [22] К 18 веку новые растения для изучения стали прибывать в Европу во все большем количестве из недавно открытых стран и европейских колоний по всему миру. В 1753 году Карл Линней опубликовал свою работу «Species Plantarum» — иерархическую классификацию видов растений, которая остаётся отправной точкой для современной ботанической номенклатуры . Это установило стандартизированную биномиальную или двухчастную схему наименования, где первое имя представляло род , а второе идентифицировало вид внутри рода. [23] В целях идентификации «Сексуальная система» Линнея разделила растения на 24 группы в зависимости от количества мужских половых органов. В 24-ю группу — Криптогамия — вошли все растения со скрытыми репродуктивными частями, мхи, печеночники, папоротники, водоросли и грибы. [24]
Расширение знаний об анатомии , морфологии и жизненных циклах растений привело к осознанию того, что между растениями существует больше естественного сходства, чем искусственная половая система Линнея. Адансон (1763), де Жюссье (1789) и Кандоль (1819) предложили различные альтернативные естественные системы классификации, которые группировали растения, используя более широкий диапазон общих признаков, и получили широкое распространение. Система Кандоля отражала его идеи о прогрессировании морфологической сложности, а более поздняя система Бентама и Хукера , которая имела влияние до середины 19 века, находилась под влиянием подхода Кандоля. Публикация Дарвином «Происхождения видов » в 1859 году и его концепция общего происхождения потребовали внесения изменений в кандольскую систему, чтобы отразить эволюционные отношения, а не просто морфологическое сходство. [25]
Большую стимуляцию ботаники оказало появление первого «современного» учебника Матиаса Шлейдена « Grundzüge der Wissenschaftlichen Botanik» , опубликованного на английском языке в 1849 году под названием «Принципы научной ботаники» . [26] Шлейден был микроскопистом и одним из первых анатомов растений, который вместе с Теодором Шванном и Рудольфом Вирховым основал клеточную теорию и был одним из первых, кто осознал значение клеточного ядра , описанного Робертом Брауном в 1831 году. [27] ] В 1855 году Адольф Фик сформулировал законы Фика , которые позволили рассчитать скорость молекулярной диффузии в биологических системах. [28]

Опираясь на генно-хромосомную теорию наследственности, выдвинутую Грегором Менделем (1822–1884), Август Вейсман (1834–1914) доказал, что наследование происходит только через гаметы . Никакие другие ячейки не могут передавать унаследованные символы. [29] Работы Кэтрин Исау (1898–1997) по анатомии растений до сих пор являются важной основой современной ботаники. Ее книги «Анатомия растений » и «Анатомия семенных растений» уже более полувека являются ключевыми учебниками по структурной биологии растений. [30] [31]

Дисциплина экологии растений была впервые разработана в конце 19 века такими ботаниками, как Евгений Варминг , который выдвинул гипотезу о том, что растения образуют сообщества , и его наставником и преемником Кристеном К. Раункером, чья система описания форм жизни растений используется до сих пор. Концепция о том, что состав растительных сообществ, таких как широколиственные леса умеренного пояса, изменяется в процессе экологической сукцессии, была разработана Генри Чандлером Коулзом , Артуром Тэнсли и Фредериком Клементсом . Клементсу приписывают идею кульминационной растительности как самой сложной растительности, которую может поддерживать окружающая среда, а Тэнсли ввел в биологию концепцию экосистем . [32] [33] [34] Опираясь на обширные более ранние работы Альфонса де Кандоля , Николай Вавилов (1887–1943) подготовил отчеты о биогеографии , центрах происхождения и истории эволюции экономических растений. [35]
В частности, с середины 1960-х годов произошли успехи в понимании физики физиологических процессов растений, таких как транспирация (перенос воды внутри тканей растений), температурная зависимость скорости испарения воды с поверхности листьев и молекулярная диффузия воды. пары и углекислый газ через устьичные отверстия. Эти разработки в сочетании с новыми методами измерения размера устьичных отверстий и скорости фотосинтеза позволили точно описать скорость газообмена между растениями и атмосферой. [36] [37] Инновации Рональда Фишера в статистическом анализе , [38] Фрэнка Йейтса и других на экспериментальной станции в Ротамстеде способствовали рациональному планированию экспериментов и анализу данных в ботанических исследованиях. [39] Открытие и идентификация растительных гормонов ауксина Кеннетом В. Тиманном в 1948 году позволило регулировать рост растений с помощью внешних химикатов. Фредерик Кэмпион Стюард был пионером в методах микроразмножения и культуры тканей растений , контролируемых растительными гормонами . [40] Синтетический ауксин 2,4-дихлорфеноксиуксусная кислота или 2,4-Д был одним из первых коммерческих синтетических гербицидов. [41]

Развитие биохимии растений в 20 веке было обусловлено современными методами органического химического анализа , такими как спектроскопия , хроматография и электрофорез . С появлением связанных молекулярно-масштабных биологических подходов молекулярной биологии , геномики , протеомики и метаболомики взаимосвязь между геномом растений и большинством аспектов биохимии, физиологии, морфологии и поведения растений может быть подвергнута детальному экспериментальному анализу. [42] Концепция, первоначально высказанная Готлибом Хаберландтом в 1902 году [43] о том, что все растительные клетки тотипотентны и их можно выращивать in vitro , в конечном итоге позволила использовать генную инженерию экспериментально, чтобы выбить ген или гены, ответственные за определенный признак, или добавьте гены, такие как GFP , которые сообщают , когда экспрессируется интересующий ген. Эти технологии позволяют биотехнологическое использование целых растений или культур растительных клеток, выращенных в биореакторах , для синтеза пестицидов , антибиотиков или других фармацевтических препаратов , а также практическое применение генетически модифицированных культур , предназначенных для повышения урожайности. [44]
Современная морфология признает континуум между основными морфологическими категориями корня, стебля (каулома), листа (филлома) и трихомы . [45] Кроме того, в нем подчеркивается структурная динамика. [46] Целью современной систематики является отражение и обнаружение филогенетических связей между растениями. [47] [48] [49] [50] Современная молекулярная филогенетика в значительной степени игнорирует морфологические признаки, полагаясь на последовательности ДНК в качестве данных. Молекулярный анализ последовательностей ДНК большинства семейств цветковых растений позволил Группе филогении покрытосеменных опубликовать в 1998 году филогению цветковых растений, ответив на многие вопросы о взаимоотношениях между семействами и видами покрытосеменных . [51] Теоретическая возможность практического метода идентификации видов и коммерческих сортов растений с помощью штрих-кодирования ДНК является предметом активных текущих исследований. [52] [53]

Изучение растений жизненно важно, поскольку они лежат в основе почти всей животной жизни на Земле, генерируя большую часть кислорода и пищи, которые обеспечивают людей и другие организмы при аэробном дыхании химической энергией, необходимой им для существования. Растения, водоросли и цианобактерии — основные группы организмов, осуществляющих фотосинтез — процесс, использующий энергию солнечного света для преобразования воды и углекислого газа [54] в сахара, которые можно использовать как в качестве источника химической энергии, так и в качестве источника органических молекул. которые используются в структурных компонентах клеток. [55] В качестве побочного продукта фотосинтеза растения выделяют в атмосферу кислород — газ, который необходим почти всем живым существам для осуществления клеточного дыхания. Кроме того, они влияют на глобальные циклы углерода и воды , а корни растений связывают и стабилизируют почву, предотвращая эрозию почвы . [56] Растения имеют решающее значение для будущего человеческого общества, поскольку они обеспечивают людей пищей, кислородом, биохимическими веществами и продуктами, а также создают и сохраняют почву. [57]
Исторически все живые существа относились либо к животным, либо к растениям [58] , а ботаника охватывала изучение всех организмов, не считавшихся животными. [59] Ботаники исследуют как внутренние функции, так и процессы внутри растительных органелл , клеток, тканей, целых растений, популяций растений и растительных сообществ. На каждом из этих уровней ботаник может заниматься классификацией ( таксономией ), филогенией и эволюцией , структурой ( анатомией и морфологией ) или функцией ( физиологией ) жизни растений. [60]
Самое строгое определение «растения» включает только «наземные растения» или эмбриофиты , к которым относятся семенные растения (голосеменные растения, включая сосны , и цветковые растения ) и криптогамы со свободными спорами, включая папоротники , плауны , печеночники , роголистники и мхи . Эмбриофиты — многоклеточные эукариоты, произошедшие от предка, который получал энергию от солнечного света путем фотосинтеза . У них есть жизненные циклы с чередованием гаплоидных и диплоидных фаз. Половая гаплоидная фаза эмбриофитов, известная как гаметофит , питает развивающийся диплоидный эмбриональный спорофит внутри своих тканей в течение, по крайней мере, части своей жизни, [61] даже у семенных растений, где сам гаметофит питается родительским спорофитом. [62] Другие группы организмов, которые ранее изучались ботаниками, включают бактерии (теперь изучаются в бактериологии ), грибы ( микология ), включая лишайниковообразующие грибы ( лихенология ), нехлорофитные водоросли ( пикология ) и вирусы ( вирусология ). Однако ботаники по-прежнему уделяют внимание этим группам, а грибы (в том числе лишайники) и фотосинтезирующие протисты обычно изучаются во вводных курсах ботаники. [63] [64]
Палеоботаники изучают древние растения в летописи окаменелостей, чтобы получить информацию об истории эволюции растений . Считается, что цианобактерии , первые фотосинтезирующие организмы, выделяющие кислород на Земле, дали начало предку растений, вступив в эндосимбиотические отношения с ранними эукариотами, в конечном итоге став хлоропластами в растительных клетках. Новые фотосинтезирующие растения (вместе со своими родственниками-водорослями) ускорили повышение содержания кислорода в атмосфере, начатое цианобактериями , изменив древнюю бескислородную восстановительную атмосферу на атмосферу, в которой свободный кислород был в изобилии на протяжении более 2 миллиардов лет. [65] [66]
Среди важных ботанических вопросов XXI века — роль растений как основных производителей в глобальном круговороте основных ингредиентов жизни: энергии, углерода, кислорода, азота и воды, а также способы, с помощью которых наше управление растениями может помочь решить глобальные экологические проблемы. управление ресурсами , сохранение , продовольственная безопасность человека , биологически инвазивные организмы , секвестрация углерода , изменение климата и устойчивость . [67]

Практически все основные продукты питания происходят либо непосредственно от первичного производства растений, либо косвенно от животных, которые их поедают. [68] Растения и другие фотосинтезирующие организмы лежат в основе большинства пищевых цепей , поскольку они используют энергию солнца и питательные вещества из почвы и атмосферы, преобразуя их в форму, которую могут использовать животные. Это то, что экологи называют первым трофическим уровнем . [69] Современные формы основных продуктов питания , таких как конопля , теф , кукуруза, рис, пшеница и другие злаковые травы, бобовые , бананы и бананы, [70] , а также конопля , лен и хлопок , выращиваемые ради волокон, являются результатом доисторического отбора на протяжении тысячелетий среди диких предковых растений с наиболее желательными характеристиками. [71]
Ботаники изучают, как растения производят пищу и как повысить урожайность, например, с помощью селекции растений , делая свою работу важной для способности человечества накормить мир и обеспечить продовольственную безопасность для будущих поколений. [72] Ботаники также изучают сорняки, которые представляют собой серьезную проблему в сельском хозяйстве, а также биологию и контроль патогенов растений в сельском хозяйстве и природных экосистемах . [73] Этноботаника – это изучение взаимоотношений между растениями и людьми. Применительно к исследованию исторических взаимоотношений растений и людей этноботаника может называться археоботаникой или палеоэтноботаникой . [74] Некоторые из самых ранних взаимоотношений между растениями и людьми возникли между коренными жителями Канады, когда они отличали съедобные растения от несъедобных. Такое отношение коренных жителей к растениям было зафиксировано этноботаниками. [75]
Биохимия растений — это изучение химических процессов, используемых растениями. Некоторые из этих процессов используются в их первичном метаболизме , например фотосинтетический цикл Кальвина и метаболизм крассуловой кислоты . [76] Другие производят специализированные материалы, такие как целлюлоза и лигнин , используемые для построения их тел, а также вторичные продукты, такие как смолы и ароматические соединения .
Растения и различные другие группы фотосинтезирующих эукариот, известных под общим названием « водоросли », имеют уникальные органеллы, известные как хлоропласты . Считается, что хлоропласты произошли от цианобактерий , которые сформировали эндосимбиотические отношения с древними предками растений и водорослей. Хлоропласты и цианобактерии содержат сине-зеленый пигмент хлорофилл а . [77] Хлорофилл a (а также его родственник хлорофилл b ) [a] поглощает свет в сине-фиолетовой и оранжево-красной частях спектра, одновременно отражая и пропуская зеленый свет, который мы видим как характерная окраска этих организмов. Энергия красного и синего света, которую поглощают эти пигменты, используется хлоропластами для создания богатых энергией соединений углерода из углекислого газа и воды посредством кислородного фотосинтеза — процесса, в ходе которого в качестве побочного продукта генерируется молекулярный кислород (O 2 ).

Энергия света, улавливаемая хлорофиллом А, первоначально имеет форму электронов (а позже и протонного градиента ), которые используются для создания молекул АТФ и НАДФН , которые временно хранят и транспортируют энергию. Их энергия используется ферментом рубиско в светонезависимых реакциях цикла Кальвина для производства молекул 3-углеродного сахара глицеральдегид-3-фосфата (G3P). Глицеральдегид-3-фосфат — первый продукт фотосинтеза и сырье, из которого синтезируются глюкоза и почти все другие органические молекулы биологического происхождения. Часть глюкозы превращается в крахмал, который хранится в хлоропластах. [81] Крахмал является характерным хранилищем энергии большинства наземных растений и водорослей, а инулин , полимер фруктозы , используется с той же целью в семействе подсолнечника Asteraceae . Некоторая часть глюкозы преобразуется в сахарозу (обычный столовый сахар) для экспорта в остальную часть растения.
В отличие от животных (у которых отсутствуют хлоропласты), растения и их родственники-эукариоты делегировали своим хлоропластам множество биохимических функций , включая синтез всех жирных кислот [82] [83] и большинства аминокислот . [84] Жирные кислоты, которые производят хлоропласты, используются для многих целей, например, в качестве материала для построения клеточных мембран и производства полимера кутина , который содержится в кутикуле растения и защищает наземные растения от высыхания. [85]
Растения синтезируют ряд уникальных полимеров , таких как молекулы полисахаридов целлюлоза , пектин и ксилоглюкан [86] , из которых построена клеточная стенка наземных растений. [87] Сосудистые наземные растения производят лигнин , полимер, используемый для укрепления вторичных клеточных стенок ксилемных трахеид и сосудов , чтобы предотвратить их разрушение, когда растение всасывает через них воду в условиях водного стресса. Лигнин также используется в других типах клеток, таких как волокна склеренхимы , которые обеспечивают структурную поддержку растения и являются основным компонентом древесины. Спорополленин — это химически стойкий полимер, обнаруженный во внешних клеточных стенках спор и пыльце наземных растений, ответственный за выживание ранних спор наземных растений и пыльцы семенных растений в летописи окаменелостей. Его широко считают маркером начала эволюции наземных растений в ордовикский период. [88] Концентрация углекислого газа в атмосфере сегодня намного ниже, чем она была, когда растения вышли на сушу в ордовикский и силурийский периоды. Многие однодольные, такие как кукуруза и ананас , а также некоторые двудольные , такие как Asteraceae , с тех пор независимо развили [89] пути, такие как метаболизм крассуловой кислоты и путь фиксации углерода C 4 для фотосинтеза, которые позволяют избежать потерь, возникающих в результате фотодыхания в более распространенном пути фиксации углерода C 3 . . Эти биохимические стратегии уникальны для наземных растений.
Фитохимия — раздел биохимии растений, изучающий главным образом химические вещества, вырабатываемые растениями в ходе вторичного метаболизма . [90] Некоторые из этих соединений являются токсинами, например, алкалоид кониин из болиголова . Другие, такие как эфирные масла , масло перечной мяты и лимонное масло, полезны благодаря своему аромату, в качестве ароматизаторов и специй (например, капсаицин ), а также в медицине в качестве фармацевтических препаратов, например, в опиуме из опиумного мака . Многие медицинские и рекреационные наркотики , такие как тетрагидроканнабинол (активный ингредиент каннабиса ), кофеин , морфин и никотин, получают непосредственно из растений. Другие представляют собой простые производные натуральных продуктов растительного происхождения. Например, обезболивающее аспирин представляет собой ацетиловый эфир салициловой кислоты , первоначально выделенный из коры ив [91] , а широкий спектр опиатных обезболивающих , таких как героин , получают путем химической модификации морфина , полученного из опийного мака . [92] Популярные стимуляторы происходят из растений, например, кофеин из кофе, чая и шоколада, а также никотин из табака. Большинство алкогольных напитков получают в результате ферментации богатых углеводами растительных продуктов, таких как ячмень (пиво), рис ( саке ) и виноград (вино). [93] Коренные американцы использовали различные растения в качестве способов лечения болезней на протяжении тысячелетий. [94] Эти знания коренных американцев о растениях были зафиксированы этноботаниками , а затем, в свою очередь, использовались фармацевтическими компаниями как способ открытия лекарств . [95]
Растения могут синтезировать цветные красители и пигменты, такие как антоцианы , отвечающие за красный цвет красного вина , желтая сварка и синяя вада , используемые вместе для производства зеленого Линкольна , индоксил , источник синего красителя индиго , традиционно используемого для окраски джинсовой ткани, и художественных пигментов гамбоджа. и поднялась еще безумнее .
Сахар, крахмал , хлопок, лен , конопля , некоторые виды веревок , древесина и древесностружечные плиты , папирус и бумага, растительные масла , воск и натуральный каучук — примеры коммерчески важных материалов, изготовленных из растительных тканей или их вторичных продуктов. Древесный уголь , чистая форма углерода, полученная пиролизом древесины, имеет долгую историю использования в качестве топлива для выплавки металлов , фильтрующего материала и адсорбента , а также материала для художников и является одним из трех ингредиентов пороха . Целлюлоза , самый распространенный в мире органический полимер, [96] может быть преобразована в энергию, топливо, материалы и химическое сырье. Продукты, изготовленные из целлюлозы, включают вискозу и целлофан , клей для обоев , биобутанол и пушечную вату . Сахарный тростник , рапс и соя относятся к растениям с высоким содержанием ферментируемых сахаров и масел, которые используются в качестве источников биотоплива , важной альтернативы ископаемому топливу , такому как биодизель . [97] Коренные американцы использовали свитграсс для защиты от насекомых, таких как комары . [98] Эти свойства сладкой травы отпугивать насекомых были позже обнаружены Американским химическим обществом в молекулах фитола и кумарина . [98]

Экология растений — это наука о функциональных отношениях между растениями и средой их обитания — средой, в которой они завершают свой жизненный цикл . Экологи растений изучают состав местных и региональных флор , их биоразнообразие , генетическое разнообразие и приспособленность , адаптацию растений к окружающей среде, а также их конкурентные или мутуалистические взаимодействия с другими видами. [100] Некоторые экологи даже полагаются на эмпирические данные коренных народов, собранные этноботаниками. [101] Эта информация может передать большой объем информации о том, какой была земля тысячи лет назад и как она изменилась за это время. [101] Цели экологии растений - понять причины их распределения, продуктивности, воздействия на окружающую среду, эволюции и реакции на изменения окружающей среды. [102]
Растения зависят от определенных эдафических (почвенных) и климатических факторов окружающей среды, но могут также изменять эти факторы. Например, они могут изменять альбедо окружающей среды , увеличивать перехват стока , стабилизировать минеральные почвы и увеличивать содержание в них органических веществ, а также влиять на местную температуру. Растения конкурируют с другими организмами в своей экосистеме за ресурсы. [103] [104] Они взаимодействуют со своими соседями в различных пространственных масштабах в группах, популяциях и сообществах , которые в совокупности составляют растительность. Регионы с характерными типами растительности и доминирующими растениями, а также схожими абиотическими и биотическими факторами, климатом и географией составляют биомы, такие как тундра или тропические леса . [105]
Травоядные животные едят растения, но растения могут защитить себя , а некоторые виды являются паразитами или даже плотоядными . Другие организмы образуют с растениями взаимовыгодные отношения. Например, микоризные грибы и ризобии снабжают растения питательными веществами в обмен на пищу, муравьиные растения привлекают муравьев для защиты, [106] медоносные пчелы , летучие мыши и другие животные опыляют цветы [107] [108] , а люди и другие животные [106 ] 109] действуют как переносчики распространения спор и семян .
Реакция растений на климатические и другие изменения окружающей среды может помочь нам понять, как эти изменения влияют на функционирование и продуктивность экосистем. Например, фенология растений может быть полезным показателем температуры в исторической климатологии , а также биологического воздействия изменения климата и глобального потепления . Палинология , анализ ископаемых отложений пыльцы в отложениях тысяч или миллионов лет назад, позволяет реконструировать климат прошлого. [110] Оценки концентрации CO 2 в атмосфере начиная с палеозоя были получены на основе плотности устьиц , формы и размеров листьев древних наземных растений . [111] Истощение озонового слоя может подвергнуть растения воздействию более высоких уровней ультрафиолетового излучения B (УФ-B), что приведет к снижению темпов роста. [112] Более того, информация, полученная в результате исследований экологии сообществ , систематики растений и таксономии , важна для понимания изменения растительности , разрушения среды обитания и исчезновения видов . [113]

Наследование у растений следует тем же фундаментальным принципам генетики, что и у других многоклеточных организмов. Грегор Мендель открыл генетические законы наследственности , изучая унаследованные черты, такие как форма, у Pisum sativum ( гороха ). То, что Мендель узнал, изучая растения, имело далеко идущие преимущества за пределами ботаники. Точно так же « прыгающие гены » были открыты Барбарой МакКлинток, когда она изучала кукурузу. [114] Тем не менее, существуют некоторые отличительные генетические различия между растениями и другими организмами.
Видовые границы у растений могут быть более слабыми, чем у животных, и часто возможны межвидовые гибриды . Известный пример — мята перечная Mentha × piperita , стерильный гибрид Mentha aquatica и мяты колосистой Mentha spicata . [115] Многие культивируемые сорта пшеницы являются результатом множественных меж- и внутривидовых скрещиваний между дикими видами и их гибридами. [116] Покрытосеменные растения с однодомными цветками часто имеют механизмы самонесовместимости , которые действуют между пыльцой и рыльцем , так что пыльца либо не достигает рыльца, либо не может прорасти и произвести мужские гаметы . [117] Это один из нескольких методов, используемых растениями для стимулирования ауткроссинга . [118] У многих наземных растений мужские и женские гаметы производятся отдельными особями. Эти виды называются раздельнополыми , когда речь идет о спорофитах сосудистых растений , и двудомными, когда речь идет о гаметофитах мохообразных . [119]
В отличие от высших животных, у которых партеногенез редок, у растений бесполое размножение может происходить по нескольким различным механизмам. Одним из примеров является образование стеблевых клубней картофеля. В частности, в арктических или альпийских местообитаниях, где возможности оплодотворения цветов животными редки, вместо цветков могут развиваться всходы или луковицы , заменяя половое размножение бесполым и создавая клональные популяции , генетически идентичные родительскому. Это один из нескольких типов апомиксиса , встречающихся у растений. Апомиксис также может возникнуть в семени , в результате которого образуется семя, содержащее эмбрион, генетически идентичный родительскому. [120]
Большинство организмов, размножающихся половым путем, диплоидны, с парными хромосомами, но удвоение числа их хромосом может происходить из-за ошибок цитокинеза . Это может произойти на ранних стадиях развития с образованием автополиплоидного или частично автополиплоидного организма, или во время нормальных процессов клеточной дифференцировки с образованием некоторых типов клеток, которые являются полиплоидными ( эндополиплоидия ), или во время образования гамет. Аллополиплоидное растение может возникнуть в результате гибридизации двух разных видов. Как автополиплоидные, так и аллополиплоидные растения часто могут нормально размножаться, но могут быть неспособны успешно скрещиваться с родительской популяцией из-за несоответствия числа хромосом. Эти растения, которые репродуктивно изолированы от родительских видов, но живут в одной и той же географической области, могут быть достаточно успешными, чтобы сформировать новый вид . [121] Некоторые в остальном стерильные полиплоиды растений все же могут размножаться вегетативно или семенным апомиксисом, образуя клональные популяции идентичных особей. [121] Твердая пшеница представляет собой плодородный тетраплоидный аллополиплоид, а мягкая пшеница представляет собой плодородный гексаплоид . Коммерческий банан является примером стерильного бессемянного триплоидного гибрида. Одуванчик обыкновенный — триплоид, дающий жизнеспособные семена апомиктическим семенем.
Как и у других эукариот, наследование эндосимбиотических органелл, таких как митохондрии и хлоропласты, у растений неменделевское . Хлоропласты наследуются от родителя-мужчины у голосеменных, но часто от родителя-женщины у цветковых растений. [122]

Значительный объем новых знаний о функциях растений получен в результате исследований молекулярной генетики модельных растений , таких как кресс-салат Тале, Arabidopsis thaliana , сорный вид семейства горчичных ( Brassicaceae ). [90] Геном или наследственная информация , содержащаяся в генах этого вида, кодируется примерно 135 миллионами пар оснований ДНК, образуя один из самых маленьких геномов среди цветковых растений . Arabidopsis был первым растением, геном которого был секвенирован в 2000 году. [123] Секвенирование некоторых других относительно небольших геномов, риса ( Oryza sativa ) [124] и Brachypodium distachyon , [125] сделало их важными модельными видами для понимания. генетика, клеточная и молекулярная биология злаков , трав и однодольных растений в целом.
Модельные растения, такие как Arabidopsis thaliana , используются для изучения молекулярной биологии растительных клеток и хлоропластов . В идеале эти организмы имеют небольшие геномы, которые хорошо известны или полностью секвенированы, небольшой размер и короткое время генерации. Кукуруза использовалась для изучения механизмов фотосинтеза и загрузки сахара во флоэму у растений C 4 . [126] Одноклеточная зеленая водоросль Chlamydomonas Reinhardtii , хотя и не является эмбриофитом , содержит зелено-пигментированный хлоропласт , родственный хлоропласту наземных растений, что делает ее полезной для изучения . [127] Красная водоросль Cyanidioschyzon merolae также использовалась для изучения некоторых основных функций хлоропластов. [128] Шпинат , [129] горох , [130] соевые бобы и мох Physcomitrella patens обычно используются для изучения биологии растительных клеток. [131]
Agrobacterium tumefaciens , почвенная ризосферная бактерия, может прикрепляться к растительным клеткам и заражать ихиндуцирующей каллус Ti-плазмидой путем горизонтального переноса генов , вызывая инфекцию каллуса, называемую болезнью корончатого галла. Шелл и Ван Монтегю (1977) предположили, что плазмида Ti может быть естественным вектором для внедрения гена Nif , ответственного за фиксацию азота в корневых клубеньках бобовых и других видов растений. [132] Сегодня генетическая модификация Ti-плазмиды является одним из основных методов внедрения трансгенов в растения и создания генетически модифицированных сельскохозяйственных культур .
Эпигенетика — это изучение наследственных изменений в функции генов , которые не могут быть объяснены изменениями в базовой последовательности ДНК [133] , но заставляют гены организма вести себя (или «выражать себя») по-другому. [134] Одним из примеров эпигенетических изменений является маркировка генов путем метилирования ДНК , которое определяет, будут ли они экспрессироваться или нет. Экспрессия генов также может контролироваться белками-репрессорами, которые прикрепляются к сайленсером областям ДНК и предотвращают экспрессию этой области кода ДНК. Эпигенетические метки могут добавляться или удаляться из ДНК на запрограммированных стадиях развития растения и отвечают, например, за различия между пыльниками, лепестками и нормальными листьями, несмотря на то, что все они имеют один и тот же основной генетический код. Эпигенетические изменения могут быть временными или сохраняться в результате последовательных клеточных делений до конца жизни клетки. Было показано, что некоторые эпигенетические изменения передаются по наследству [135] , в то время как другие восстанавливаются в зародышевых клетках.
Эпигенетические изменения в биологии эукариот служат регуляции процесса клеточной дифференцировки . В ходе морфогенеза тотипотентные стволовые клетки превращаются в различные плюрипотентные клеточные линии эмбриона , которые , в свою очередь , становятся полностью дифференцированными клетками. Одна оплодотворенная яйцеклетка, зигота , по мере продолжения деления дает начало множеству различных типов растительных клеток , включая паренхиму , элементы сосудов ксилемы , ситовидные трубки флоэмы, замыкающие клетки эпидермиса и т. д . Этот процесс является результатом эпигенетической активации одних генов и ингибирования других. [136]
В отличие от животных, многие растительные клетки, особенно клетки паренхимы , не дифференцируются окончательно, оставаясь тотипотентными и способными давать начало новому отдельному растению. Исключения составляют сильно лигнифицированные клетки, склеренхима и ксилема, погибающие при созревании, а также ситовидные трубки флоэмы, в которых отсутствуют ядра. Хотя растения используют многие из тех же эпигенетических механизмов, что и животные, например ремоделирование хроматина , альтернативная гипотеза заключается в том, что растения устанавливают свои паттерны экспрессии генов, используя позиционную информацию из окружающей среды и окружающих клеток, чтобы определить судьбу своего развития. [137]
Эпигенетические изменения могут привести к парамутациям , которые не соответствуют правилам менделевского наследия. Эти эпигенетические метки передаются от одного поколения к другому, при этом один аллель вызывает изменение другого. [138]

Хлоропласты растений имеют ряд биохимических, структурных и генетических сходств с цианобактериями (широко, но ошибочно известными как «сине-зеленые водоросли») и, как полагают, произошли в результате древних эндосимбиотических отношений между предковой эукариотической клеткой и резидентом цианобактерии. . [139] [140] [141] [142]
Водоросли представляют собой полифилетическую группу и относятся к различным отделам, некоторые из которых более тесно связаны с растениями, чем другие . Между ними существует множество различий по таким характеристикам, как состав клеточной стенки, биохимия, пигментация, структура хлоропластов и запасы питательных веществ. Отделение водорослей Charophyta , сестра отдела зеленых водорослей Chlorophyta , считается предком настоящих растений. [143] Класс Charophyte Charophyceae и подцарство наземных растений Embryophyta вместе образуют монофилетическую группу или кладу Streptophytina . [144]
Несосудистые наземные растения представляют собой эмбриофиты , у которых отсутствуют сосудистые ткани ксилемы и флоэмы . К ним относятся мхи , печеночники и роголистники . Птеридофитные сосудистые растения с настоящей ксилемой и флоэмой, которые размножаются спорами, прорастающими в свободноживущие гаметофиты, эволюционировали в силурийский период и разделились на несколько линий в позднем силуре и раннем девоне . Представители плаунов дожили до наших дней. К концу девонского периода несколько групп, в том числе плауноногие , сфенофиллы и прогимноспермы , независимо развили «мегаспорию» — их споры имели два разных размера: более крупные мегаспоры и меньшие микроспоры. Их редуцированные гаметофиты развиваются из мегаспор, остающихся в спорообразующих органах (мегаспорангиях) спорофита, состояние, известное как эндоспория. Семена состоят из эндоспорового мегаспорангия, окруженного одним или двумя покровными слоями ( интегументами ). Молодой спорофит развивается внутри семени, которое при прорастании расщепляется, высвобождая его. Самые ранние известные семенные растения относятся к позднему девонскому фаменскому этапу. [145] [146] После эволюции семенного образа жизни семенные растения диверсифицировались, дав начало ряду ныне вымерших групп, включая семенные папоротники , а также современные голосеменные и покрытосеменные растения. [147] Голосеменные растения производят «голые семена», не полностью заключенные в завязь; современные представители — хвойные , саговники , гинкго , гнеталесы . Покрытосеменные производят семена, заключенные в такую структуру, как плодолистик или завязь . [148] [149] Продолжающиеся исследования молекулярной филогенетики живых растений, по-видимому, показывают, что покрытосеменные растения являются сестринскими кладами голосеменных растений. [150]

Физиология растений охватывает всю внутреннюю химическую и физическую деятельность растений, связанную с жизнью. [151] Химические вещества, полученные из воздуха, почвы и воды, составляют основу всего метаболизма растений . Энергия солнечного света, улавливаемая в результате кислородного фотосинтеза и выделяемая при клеточном дыхании , является основой практически всей жизни. Фотоавтотрофы , включая все зеленые растения, водоросли и цианобактерии , собирают энергию непосредственно из солнечного света путем фотосинтеза. Гетеротрофы , включая всех животных, все грибы, все полностью паразитические растения и нефотосинтезирующие бактерии, поглощают органические молекулы, вырабатываемые фотоавтотрофами, и дышат ими или используют их в построении клеток и тканей. [152] Дыхание — это окисление соединений углерода путем расщепления их на более простые структуры с высвобождением содержащейся в них энергии, что по существу противоположно фотосинтезу. [153]
Молекулы перемещаются внутри растений посредством транспортных процессов, которые происходят в различных пространственных масштабах . Субклеточный транспорт ионов, электронов и молекул, таких как вода и ферменты, происходит через клеточные мембраны . Минералы и вода переносятся от корней к другим частям растения с транспирационным потоком . Диффузия , осмос , активный транспорт и массовый поток — все это различные способы транспорта. [154] Примерами элементов, которые растениям необходимо транспортировать, являются азот , фосфор , калий , кальций , магний и сера . У сосудистых растений эти элементы извлекаются из почвы в виде растворимых ионов корнями и транспортируются по растению в ксилеме. Большая часть элементов, необходимых для питания растений , поступает в результате химического распада минералов почвы. [155] Сахароза , образующаяся в результате фотосинтеза, транспортируется из листьев в другие части растения по флоэме, а растительные гормоны переносятся посредством множества процессов.

Растения не пассивны, а реагируют на внешние сигналы , такие как свет, прикосновение и травмы, перемещаясь или растущих к раздражителю или от него, в зависимости от обстоятельств. Осязаемыми свидетельствами сенсорной чувствительности являются почти мгновенное свертывание листочков Mimosa pudica , ловушки для насекомых венериной мухоловки и пузырчатки , а также поллинии орхидей. [157]
Гипотеза о том, что рост и развитие растений координируется растительными гормонами или регуляторами роста растений, впервые возникла в конце 19 века. Дарвин экспериментировал с движением побегов и корней растений в направлении света [158] и силы тяжести и пришел к выводу: «Едва ли будет преувеличением сказать, что кончик корешка... действует как мозг одного из низших животных... направляя движение несколько движений». [159] Примерно в то же время роль ауксинов (от греческого auxein — расти) в контроле роста растений была впервые описана голландским учёным Фрицем Вентом . [160] Первый известный ауксин, индол-3-уксусная кислота (ИУК), которая способствует росту клеток, был выделен из растений только примерно 50 лет спустя. [161] Это соединение опосредует тропическую реакцию побегов и корней на свет и силу тяжести. [162] Открытие в 1939 году того, что каллус растений можно поддерживать в культуре, содержащей ИУК, а затем в 1947 году наблюдение о том, что его можно стимулировать к образованию корней и побегов путем контроля концентрации гормонов роста, стали ключевыми шагами в развитии биотехнологии растений. и генетическая модификация. [163]
Цитокинины — это класс растительных гормонов, названных в честь их контроля деления клеток (особенно цитокинеза ). Природный цитокинин зеатин был обнаружен в кукурузе Zea mays и является производным пурина аденина . Зеатин вырабатывается в корнях и транспортируется в побеги в ксилеме, где способствует делению клеток, развитию почек и зеленению хлоропластов. [164] [165] Гибберелины , такие как гибберелиновая кислота, представляют собой дитерпены , синтезируемые из ацетил-КоА по мевалонатному пути . Они участвуют в прорастании и нарушении покоя семян, в регулировании высоты растений путем контроля удлинения стебля и контроля цветения. [166] Абсцизовая кислота (АБК) встречается во всех наземных растениях, кроме печеночников, и синтезируется из каротиноидов в хлоропластах и других пластидах. Он ингибирует деление клеток, способствует созреванию и покою семян, а также способствует закрытию устьиц. Он был назван так потому, что первоначально считалось, что он контролирует отпадение . [167] Этилен — газообразный гормон, который вырабатывается во всех тканях высших растений из метионина . Сейчас известно, что это гормон, который стимулирует или регулирует созревание и опадение плодов, [168] [169] и он, или синтетический регулятор роста этефон , который быстро метаболизируется с образованием этилена, используются в промышленных масштабах для ускорения созревания хлопка. , ананасы и другие климактерические культуры.
Другим классом фитогормонов являются жасмонаты , впервые выделенные из масла Jasminum grandiflorum [170] , которые регулируют реакцию на раны у растений, разблокируя экспрессию генов, необходимых для системной приобретенной реакции устойчивости к атаке патогенов. [171]
Помимо того, что свет является основным источником энергии для растений, он действует как сигнальное устройство, предоставляя растению информацию, например, сколько солнечного света растение получает каждый день. Это может привести к адаптивным изменениям в процессе, известном как фотоморфогенез . Фитохромы — фоторецепторы растений, чувствительные к свету. [172]
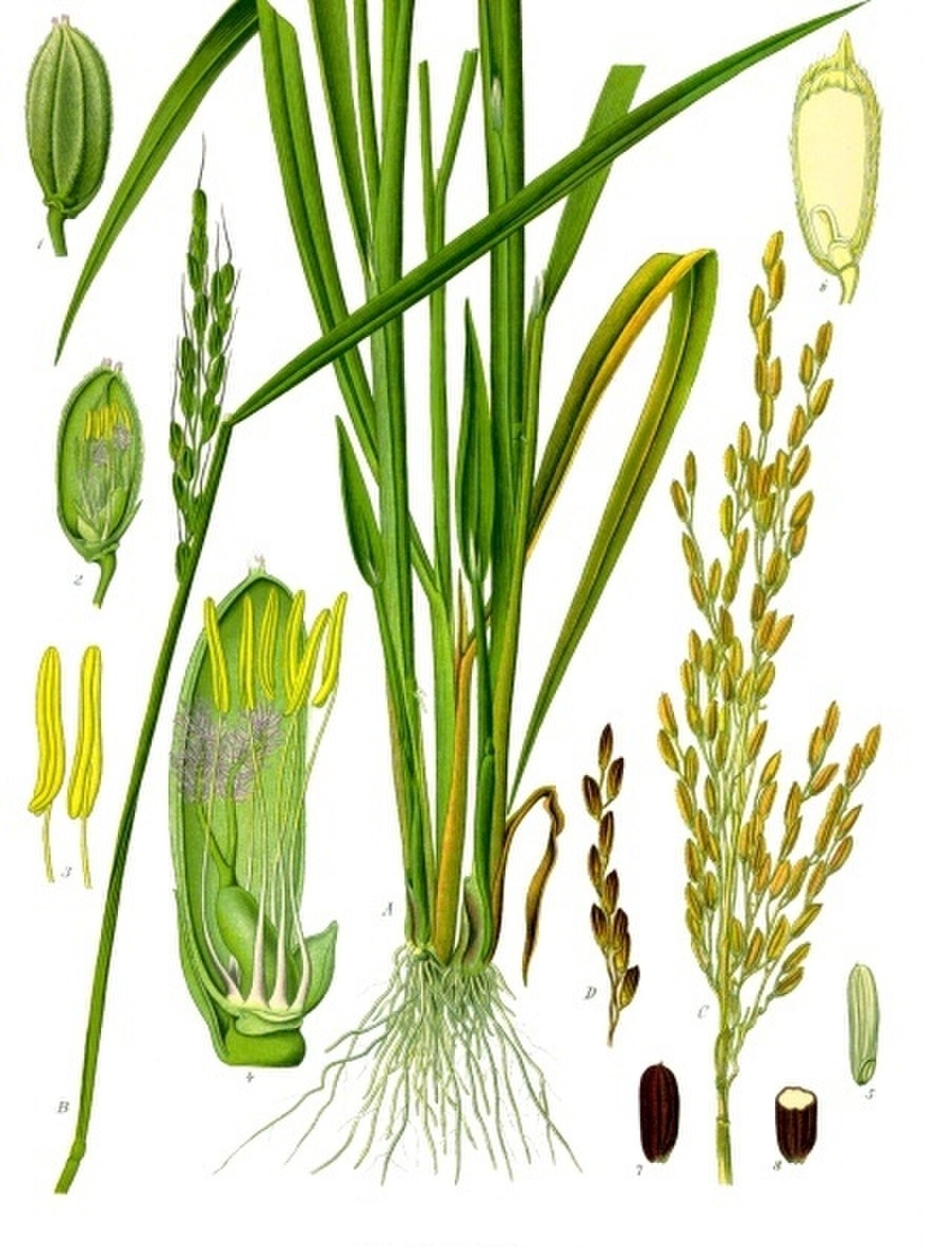
Анатомия растений — это изучение строения растительных клеток и тканей, тогда как морфология растений — это изучение их внешней формы. [173] Все растения являются многоклеточными эукариотами, их ДНК хранится в ядрах. [174] [175] Характерные особенности растительных клеток , которые отличают их от клеток животных и грибов, включают первичную клеточную стенку , состоящую из полисахаридов целлюлозы , гемицеллюлозы и пектина , [176] более крупные вакуоли , чем в животных клетках, и наличие пластид. с уникальными фотосинтетическими и биосинтетическими функциями, как у хлоропластов. Другие пластиды содержат продукты хранения, такие как крахмал ( амилопласты ) или липиды ( элайопласты ). Уникально то, что клетки стрептофитов и клетки зеленых водорослей отряда Trentepohliales [177] делятся путем построения фрагмопласта в качестве матрицы для построения клеточной пластинки на поздних стадиях клеточного деления . [81]
![Схема «типичного» эвдикота, наиболее распространенного вида растения (три пятых всех видов растений).[178] Однако ни одно растение на самом деле не выглядит именно так.](http://upload.wikimedia.org/wikipedia/commons/thumb/a/a9/Plant.svg/1280px-Plant.svg.png)
Тела сосудистых растений, включая плауны , папоротники и семенные растения ( голосеменные и покрытосеменные ), обычно имеют надземную и подземную подсистемы. Побеги состоят из стеблей с зелеными фотосинтезирующими листьями и репродуктивными структурами. Подземные васкуляризированные корни несут на кончиках корневые волоски и обычно лишены хлорофилла. [179] Несосудистые растения, печеночники , роголистники и мхи не образуют проникающих в землю сосудистых корней, и большая часть растений участвует в фотосинтезе. [180] Поколение спорофитов у печеночников не является фотосинтезирующим, но может частично удовлетворять свои энергетические потребности за счет фотосинтеза у мхов и роголистников . [181]
Корневая система и система побегов взаимозависимы: обычно нефотосинтетическая корневая система зависит от побеговой системы для питания, а обычно фотосинтетическая побеговая система зависит от воды и минералов корневой системы. [179] Клетки каждой системы способны создавать клетки другой и производить дополнительные побеги или корни. [182] Столоны и клубни являются примерами побегов, у которых могут образовываться корни. [183] Корни, которые распространяются близко к поверхности, например, у ив, могут давать побеги и, в конечном итоге, новые растения. [184] В случае потери одной из систем другая часто может восстановить ее. Фактически, из одного листа можно вырастить целое растение, как в случае с растениями секты Streptocarpus . Сенполия , [185] или даже одна клетка , которая может дедифференцироваться в каллус (массу неспециализированных клеток), из которого может вырасти новое растение. [182] У сосудистых растений ксилема и флоэма являются проводящими тканями, которые транспортируют ресурсы между побегами и корнями. Корни часто приспособлены для хранения пищевых продуктов, таких как сахар или крахмал , [179] как у сахарной свеклы и моркови. [184]
Стебли в основном обеспечивают поддержку листьям и репродуктивным структурам, но могут хранить воду в суккулентных растениях, таких как кактусы , пищу, как в клубнях картофеля , или размножаться вегетативно, как в столонах растений клубники , или в процессе отводков . [186] Листья собирают солнечный свет и осуществляют фотосинтез . [187] Большие, плоские, гибкие, зеленые листья называются лиственными листьями. [188] Голосеменные растения , такие как хвойные деревья , саговники , гинкго и гнетофиты , представляют собой семенные растения с открытыми семенами. [189] Покрытосеменные растения — это семенные растения , которые дают цветы и имеют закрытые семена. [148] Древесные растения, такие как азалии и дубы , проходят вторичную фазу роста, в результате которой образуются два дополнительных типа тканей: древесина (вторичная ксилема ) и кора (вторичная флоэма и пробка ). Все голосеменные и многие покрытосеменные растения являются древесными растениями. [190] Некоторые растения размножаются половым путем, некоторые бесполым, а некоторые - обоими способами. [191]
Хотя ссылки на основные морфологические категории, такие как корень, стебель, лист и трихома, полезны, следует иметь в виду, что эти категории связаны через промежуточные формы, так что в результате получается континуум между категориями. [192] Более того, структуры можно рассматривать как процессы, то есть комбинации процессов. [46]

Систематическая ботаника является частью систематической биологии, которая занимается ареалом и разнообразием организмов и их взаимоотношениями, особенно определяемыми их эволюционной историей. [193] Оно включает или связано с биологической классификацией, научной таксономией и филогенетикой . Биологическая классификация — это метод, с помощью которого ботаники группируют организмы в такие категории, как роды или виды . Биологическая классификация — это форма научной систематики . Современная систематика уходит корнями в работы Карла Линнея , который сгруппировал виды по общим физическим характеристикам. С тех пор эти группировки были пересмотрены, чтобы лучше соответствовать дарвиновскому принципу общего происхождения – группировке организмов по происхождению, а не по поверхностным характеристикам . Хотя ученые не всегда сходятся во мнении относительно того, как классифицировать организмы, молекулярная филогенетика , которая использует последовательности ДНК в качестве данных, привела к множеству недавних пересмотров эволюционных направлений и, вероятно, будет продолжать это делать. Доминирующая система классификации называется таксономией Линнея . Он включает ранги и биномиальную номенклатуру . Номенклатура ботанических организмов кодифицирована в Международном кодексе номенклатуры водорослей, грибов и растений (ICN) и администрируется Международным ботаническим конгрессом . [194] [195]
Королевство Plantae принадлежит к домену Eukaryota и рекурсивно разбивается до тех пор, пока каждый вид не будет классифицирован отдельно. Порядок такой: Королевство ; Тип (или отдел); Сорт ; Заказ ; Семья ; Род (множественное число родов ); Разновидность . Научное название растения представляет его род и виды внутри рода, в результате чего для каждого организма создается единое всемирное название. [195] Например, тигровая лилия — Lilium columbianum . Lilium — это род, а columbianum — видовой эпитет . Сочетание и есть название вида. При написании научного названия организма первую букву рода следует писать с заглавной буквы, а весь видовой эпитет писать строчными. Кроме того, весь термин обычно выделяется курсивом (или подчеркивается, если курсив недоступен). [196] [197] [198]
Эволюционные связи и наследственность группы организмов называют ее филогенией . Филогенетические исследования пытаются обнаружить филогении. Основной подход заключается в использовании сходств на основе общего наследования для определения отношений. [199] Например, виды перескии представляют собой деревья или кусты с выступающими листьями. Они явно не похожи на типичный безлистный кактус , такой как эхинокактус . Однако и Pereskia , и Echinocactus имеют шипы, образующиеся из ареол (узкоспециализированных подушечек), что позволяет предположить, что эти два рода действительно связаны. [200] [201]
Оценка отношений на основе общих признаков требует осторожности, поскольку растения могут походить друг на друга в результате конвергентной эволюции , в которой признаки возникли независимо. У некоторых молочайов безлистные округлые тела, приспособленные к сохранению воды, подобные телам шаровидных кактусов, но такие особенности, как структура их цветков, ясно показывают, что эти две группы не являются тесно связанными. Кладистический метод использует систематический подход к признакам, различая те, которые не несут никакой информации об общей эволюционной истории – например, те, которые развивались отдельно в разных группах ( гомоплазии ) или оставшиеся от предков ( плезиоморфии ) – и производные признаки, которые были передающиеся от нововведений от общего предка ( апоморфии ). Только производные признаки, такие как ареолы кактусов, образующие шипы, служат доказательством происхождения от общего предка. Результаты кладистического анализа выражаются в виде кладограмм : древовидных диаграмм, показывающих закономерности эволюционного ветвления и происхождения. [202]
С 1990-х годов преобладающим подходом к построению филогении живых растений была молекулярная филогенетика , которая использует молекулярные признаки, особенно последовательности ДНК , а не морфологические признаки, такие как наличие или отсутствие шипов и ареол. Разница в том, что для определения эволюционных отношений используется сам генетический код, а не косвенно, через признаки, которые он порождает. Клайв Стейс описывает это как «прямой доступ к генетической основе эволюции». [203] В качестве простого примера: до использования генетических данных считалось, что грибы либо являются растениями, либо более тесно связаны с растениями, чем с животными. Генетические данные свидетельствуют о том, что истинные эволюционные взаимоотношения многоклеточных организмов показаны на кладограмме ниже: грибы более тесно связаны с животными, чем с растениями. [204]
В 1998 году Группа филогении покрытосеменных опубликовала филогению цветковых растений, основанную на анализе последовательностей ДНК большинства семейств цветковых растений. В результате этой работы теперь получены ответы на многие вопросы, например, какие семейства представляют самые ранние ветви покрытосеменных . [51] Исследование того, как виды растений связаны друг с другом, позволяет ботаникам лучше понять процесс эволюции растений. [205] Несмотря на изучение модельных растений и растущее использование данных ДНК, среди систематиков продолжается работа и дискуссии о том, как лучше всего классифицировать растения по различным таксонам . [206] Технологические разработки, такие как компьютеры и электронные микроскопы, значительно повысили уровень детализации изучаемых данных и скорость анализа данных. [207]
В ботанике в настоящее время используются несколько символов. Ряд других устарели; например, Линней использовал планетарные символы ⟨♂⟩ (Марс) для двулетних растений, ⟨♃⟩ (Юпитер) для травянистых многолетних растений и ⟨♄⟩ (Сатурн) для древесных многолетних растений, исходя из периодов обращения планет 2, 12 и 30. годы; и Уиллд использовал ⟨♄⟩ (Сатурн) для среднего рода в дополнение к ⟨☿⟩ (Меркурий) для гермафродита. [208] По-прежнему используются следующие символы: [209]
{{cite book}}: |journal=игнорируется ( помощь )






{kind=link}
{kind=link}
{kind=link}