Глюкоза — это сахар с молекулярной формулой C6H12O6 . Глюкоза — это в целом самый распространенный моносахарид , [4] подкатегория углеводов . Глюкоза в основном производится растениями и большинством водорослей в процессе фотосинтеза из воды и углекислого газа с использованием энергии солнечного света. Глюкоза используется растениями для производства целлюлозы — самого распространенного углевода в мире — для использования в клеточных стенках , а также всеми живыми организмами для производства аденозинтрифосфата (АТФ), который используется клеткой в качестве энергии. [5] [6] [7]
В энергетическом обмене веществ глюкоза является важнейшим источником энергии во всех организмах . Глюкоза для метаболизма хранится в виде полимера , в растениях в основном в виде амилозы и амилопектина , а у животных в виде гликогена . Глюкоза циркулирует в крови животных в виде сахара в крови . [5] [7] Естественной формой глюкозы является d -глюкоза, в то время как ее стереоизомер l -глюкоза производится синтетически в сравнительно небольших количествах и является менее биологически активным. [7] Глюкоза представляет собой моносахарид, содержащий шесть атомов углерода и альдегидную группу, и, следовательно, является альдогексозой . Молекула глюкозы может существовать как в форме открытой цепи (ациклической), так и в форме кольца (циклической). Глюкоза встречается в природе и находится в свободном состоянии во фруктах и других частях растений. У животных глюкоза высвобождается при распаде гликогена в процессе, известном как гликогенолиз .
Глюкоза, как внутривенный раствор сахара , входит в список основных лекарственных средств Всемирной организации здравоохранения . [8] Она также входит в список в сочетании с хлоридом натрия (поваренной солью). [8]
Название «глюкоза» происходит от древнегреческого γλεῦκος ( gleûkos ) «вино, сусло», от γλυκύς ( glykýs ) «сладкий». [9] [10] Суффикс -ose является химическим классификатором, обозначающим сахар.
Глюкоза была впервые выделена из изюма в 1747 году немецким химиком Андреасом Маргграфом . [11] [12] Глюкоза была обнаружена в винограде другим немецким химиком – Иоганном Тобиасом Ловицем – в 1792 году и была выделена как отличная от тростникового сахара ( сахарозы ). Глюкоза – это термин, введенный Жаном Батистом Дюма в 1838 году, который преобладает в химической литературе. Фридрих Август Кекуле предложил термин декстроза (от латинского dexter , что означает «правый»), потому что в водном растворе глюкозы плоскость линейно поляризованного света повернута вправо. Напротив, l-фруктоза (обычно называемая d- фруктозой) (кетогексозой) и l-глюкоза ( l -глюкоза) поворачивают линейно поляризованный свет влево. Более ранняя нотация, соответствующая вращению плоскости линейно поляризованного света ( d и l -номенклатура), позднее была заменена на d- и l -нотацию , которая относится к абсолютной конфигурации асимметричного центра, наиболее удаленного от карбонильной группы, и соответствует конфигурации d- или l -глицеральдегида. [13] [14]
Поскольку глюкоза является основной потребностью многих организмов, правильное понимание ее химического состава и структуры внесло большой вклад в общее развитие органической химии . Это понимание произошло в значительной степени в результате исследований Эмиля Фишера , немецкого химика, который получил Нобелевскую премию по химии 1902 года за свои открытия. [15] Синтез глюкозы установил структуру органического материала и, следовательно, сформировал первое окончательное подтверждение теорий Якоба Генрикуса Вант 'т Гоффа о химической кинетике и расположении химических связей в углеродсодержащих молекулах. [16] Между 1891 и 1894 годами Фишер установил стереохимическую конфигурацию всех известных сахаров и правильно предсказал возможные изомеры , применив теорию Вант 'т Гоффа об асимметричных атомах углерода. Названия изначально относились к природным веществам. Их энантиомеры получили одинаковые названия с введением систематических номенклатур, учитывающих абсолютную стереохимию (например, номенклатура Фишера, номенклатура d / l ).
За открытие метаболизма глюкозы Отто Мейерхоф получил Нобелевскую премию по физиологии и медицине в 1922 году. [17] Ганс фон Эйлер-Хельпин был удостоен Нобелевской премии по химии вместе с Артуром Харденом в 1929 году за их «исследования ферментации сахара и их доли ферментов в этом процессе». [18] [19] В 1947 году Бернардо Усай (за открытие роли гипофиза в метаболизме глюкозы и производных углеводов), а также Карл и Герти Кори (за открытие превращения гликогена из глюкозы) получили Нобелевскую премию по физиологии и медицине. [20] [21] [22] В 1970 году Луис Лелуар был удостоен Нобелевской премии по химии за открытие производных глюкозы сахарных нуклеотидов в биосинтезе углеводов. [23]
Глюкоза образует белые или бесцветные твердые вещества, которые хорошо растворимы в воде и уксусной кислоте , но плохо растворимы в метаноле и этаноле . Они плавятся при 146 °C (295 °F) ( α ) и 150 °C (302 °F) ( beta ), разлагаются , начиная с 188 °C (370 °F) с выделением различных летучих продуктов, в конечном итоге оставляя остаток углерода . [24] Глюкоза имеет значение pKa 12,16 при 25 °C (77 °F) в воде. [25]
С шестью атомами углерода, он классифицируется как гексоза , подкатегория моносахаридов . d - Глюкоза является одним из шестнадцати стереоизомеров альдогексозы . d - Изомер , d -глюкоза, также известный как декстроза, широко встречается в природе, но l -изомер, l -глюкоза , не встречается. Глюкозу можно получить путем гидролиза углеводов, таких как молочный сахар ( лактоза ), тростниковый сахар (сахароза), мальтоза , целлюлоза , гликоген и т. д. Декстрозу обычно производят в коммерческих целях из крахмалов , таких как кукурузный крахмал в США и Японии, из картофельного и пшеничного крахмала в Европе и из крахмала тапиоки в тропических районах. [26] В процессе производства используется гидролиз посредством пропаривания под давлением при контролируемом pH в струе с последующей ферментативной деполимеризацией. [27] Несвязанная глюкоза является одним из основных ингредиентов меда . [28] [29] [30] [31] [32]
Термин «декстроза» часто используется в клиническом (связанном с состоянием здоровья пациента) или пищевом контексте (связанном с диетическим питанием, например, на этикетках продуктов питания или в рекомендациях по питанию), в то время как «глюкоза» используется в биологическом или физиологическом контексте (химические процессы и молекулярные взаимодействия), [33] [34] [35] [36], но оба термина относятся к одной и той же молекуле, а именно к D-глюкозе. [35] [37]
Моногидрат декстрозы — это гидратированная форма D-глюкозы, то есть это молекула глюкозы с присоединенной дополнительной молекулой воды. [ 38] Его химическая формула — C 6 H 12 O 6 · H 2 O. [38] [39] Моногидрат декстрозы также называется гидратированной D-глюкозой и обычно производится из растительных крахмалов. [38] [40] Моногидрат декстрозы используется как преобладающий тип декстрозы в пищевых продуктах, таких как смеси для напитков — это распространенная форма глюкозы, широко используемая в качестве пищевой добавки при производстве продуктов питания. Моногидрат декстрозы в основном потребляется в Северной Америке в виде кукурузного сиропа или кукурузного сиропа с высоким содержанием фруктозы . [35]
Безводная декстроза , с другой стороны, представляет собой глюкозу, к которой не присоединены молекулы воды. [40] [41] Безводные химические вещества обычно производятся путем удаления воды из гидратированного вещества с помощью таких методов, как нагревание или высушивание (десикация). [42] [43] [44] Моногидрат декстрозы может быть дегидратирован до безводной декстрозы в промышленных условиях. [45] [46] Моногидрат декстрозы состоит приблизительно из 9,5% воды по массе; в процессе дегидратации это содержание воды устраняется, чтобы получить безводную (сухую) декстрозу. [40]
Безводная декстроза имеет химическую формулу C6H12O6 , без какой - либо присоединенной молекулы воды, что соответствует формуле глюкозы. [ 38] Безводная декстроза на открытом воздухе имеет тенденцию впитывать влагу и превращаться в моногидрат, и ее производство обходится дороже. [40] Безводная декстроза (безводная D-глюкоза) имеет повышенную стабильность и увеличенный срок хранения, [43] имеет медицинское применение, например, в оральном тесте на толерантность к глюкозе . [47]
В то время как молекулярная масса (молярная масса) моногидрата D-глюкозы составляет 198,17 г/моль, [48] [49] то для безводной D-глюкозы она составляет 180,16 г/моль [50] [51] [52] Плотность этих двух форм глюкозы также различна. [ указать ]
С точки зрения химической структуры глюкоза является моносахаридом, то есть простым сахаром. Глюкоза содержит шесть атомов углерода и альдегидную группу , и поэтому является альдогексозой . Молекула глюкозы может существовать как в форме открытой цепи (ациклической), так и в форме кольца (циклической) — из-за наличия спиртовых и альдегидных или кетонных функциональных групп форма с прямой цепью может легко преобразоваться в подобную стулу полуацетальную кольцевую структуру, обычно встречающуюся в углеводах. [53]

Глюкоза присутствует в твердой форме в виде моногидрата с замкнутым пирановым кольцом (моногидрат α-глюкопиранозы, иногда менее точно известный как гидрат декстрозы). В водном растворе, с другой стороны, она в небольшой степени является открытой цепью и присутствует преимущественно в виде α- или β- пиранозы , которые взаимопревращаются. Из водных растворов могут быть кристаллизованы три известные формы: α-глюкопираноза, β-глюкопираноза и моногидрат α-глюкопиранозы. [54] Глюкоза является строительным блоком дисахаридов лактозы и сахарозы (тростникового или свекловичного сахара), олигосахаридов, таких как раффиноза , и полисахаридов, таких как крахмал , амилопектин , гликоген и целлюлоза . [7] [55] Температура стеклования глюкозы составляет 31 °C (88 °F), а константа Гордона-Тейлора (экспериментально определенная константа для прогнозирования температуры стеклования для различных массовых долей смеси двух веществ) [55] составляет 4,5. [56]

Форма глюкозы с открытой цепью составляет менее 0,02% молекул глюкозы в водном растворе при равновесии. [57] Остальное — одна из двух циклических полуацетальных форм. В форме с открытой цепью молекула глюкозы имеет открытую (в отличие от циклической ) неразветвленную основу из шести атомов углерода, где C-1 является частью альдегидной группы H(C=O)− . Поэтому глюкозу также классифицируют как альдозу или альдогексозу . Альдегидная группа делает глюкозу восстанавливающим сахаром, что дает положительную реакцию с тестом Фелинга .
В растворах форма глюкозы с открытой цепью (либо « D- », либо « L- ») существует в равновесии с несколькими циклическими изомерами , каждый из которых содержит кольцо из атомов углерода, замкнутое одним атомом кислорода. Однако в водном растворе более 99% молекул глюкозы существуют в виде пиранозных форм. Форма с открытой цепью ограничена примерно 0,25%, а формы фуранозы существуют в незначительных количествах. Термины «глюкоза» и « D -глюкоза» обычно используются также для этих циклических форм. Кольцо возникает из формы с открытой цепью в результате внутримолекулярной реакции нуклеофильного присоединения между альдегидной группой (в положении C-1) и гидроксильной группой либо C-4, либо C-5, образуя полуацетальную связь, −C(OH)H−O− .
Реакция между C-1 и C-5 дает шестичленную гетероциклическую систему, называемую пиранозой, которая представляет собой моносахаридный сахар (отсюда «-оза»), содержащий дериватизированный скелет пирана . (Гораздо более редкая) реакция между C-1 и C-4 дает пятичленное фуранозное кольцо, названное в честь циклического эфира фурана . В любом случае каждый углерод в кольце имеет один водород и один присоединенный гидроксил, за исключением последнего углерода (C-4 или C-5), где гидроксил заменен остатком открытой молекулы (который является −(C(CH 2 OH)HOH)−H или −(CHOH)−H соответственно).
Реакция замыкания кольца может дать два продукта, обозначенных «α-» и «β-». Когда молекула глюкопиранозы изображена в проекции Хаворта , обозначение «α-» означает, что гидроксильная группа, присоединенная к C-1, и группа −CH 2 OH при C-5 находятся на противоположных сторонах плоскости кольца ( транс -расположение), тогда как «β-» означает, что они находятся на одной стороне плоскости ( цис -расположение). Таким образом, изомер D -глюкозы с открытой цепью дает четыре различных циклических изомера: α- D -глюкопиранозу, β- D -глюкопиранозу, α- D -глюкофуранозу и β- D -глюкофуранозу. Эти пять структур существуют в равновесии и взаимопревращаются, и взаимопревращение происходит гораздо быстрее при кислотном катализе .

Другой изомер с открытой цепью L -глюкоза аналогичным образом дает начало четырем различным циклическим формам L -глюкозы, каждая из которых является зеркальным отражением соответствующей D -глюкозы.
Кольцо глюкопиранозы (α или β) может принимать несколько неплоских форм, аналогичных конформациям «кресло» и «лодка» циклогексана . Аналогично, кольцо глюкофуранозы может принимать несколько форм, аналогичных конформациям «конверт» циклопентана .
В твердом состоянии наблюдаются только глюкопиранозные формы.
Некоторые производные глюкофуранозы, такие как 1,2-O-изопропилиден-D-глюкофураноза, стабильны и могут быть получены в чистом виде в виде кристаллических твердых веществ. [58] [59] Например, реакция α-D-глюкозы с пара-толилбороновой кислотой H 3 C−(C 6 H 4 )−B(OH) 2 преобразует нормальное пиранозное кольцо с образованием 4-кратного эфира α-D-глюкофураноза-1,2:3,5-бис( п -толилборонат). [60]
Мутаротация состоит из временного обращения реакции формирования кольца, что приводит к форме с открытой цепью, за которой следует реформирование кольца. Шаг замыкания кольца может использовать другую группу −OH, чем та, которая была воссоздана шагом открытия (таким образом, переключение между пиранозной и фуранозной формами), или новая полуацетальная группа, созданная на C-1, может иметь ту же или противоположную направленность, что и исходная (таким образом, переключение между α- и β-формами). Таким образом, хотя форма с открытой цепью едва обнаруживается в растворе, она является существенным компонентом равновесия.
Форма с открытой цепью термодинамически нестабильна и спонтанно изомеризуется в циклические формы. (Хотя реакция замыкания кольца теоретически могла бы создавать кольца из четырех или трех атомов, они были бы сильно напряжены и не наблюдаются на практике.) В растворах при комнатной температуре четыре циклических изомера взаимопревращаются в течение времени в часах в процессе, называемом мутаротацией . [61] Начиная с любых пропорций, смесь сходится к стабильному соотношению α:β 36:64. Соотношение было бы α:β 11:89, если бы не влияние аномерного эффекта . [62] Мутаротация происходит значительно медленнее при температурах, близких к 0 °C (32 °F).
Будь то в воде или в твердой форме, d -(+)-глюкоза является правовращающей , то есть она будет вращать направление поляризованного света по часовой стрелке, если смотреть на источник света. Эффект обусловлен хиральностью молекул , и действительно, зеркальный изомер, l -(−)-глюкоза, является левовращающей (вращает поляризованный свет против часовой стрелки) на ту же величину. Сила эффекта различна для каждого из пяти таутомеров .
Префикс d - не относится напрямую к оптическим свойствам соединения. Он указывает на то, что хиральный центр C-5 имеет ту же ручность, что и d -глицеральдегид (который был так обозначен, потому что является правовращающим). Тот факт, что d -глюкоза является правовращающей, является совокупным эффектом ее четырех хиральных центров, а не только C-5; некоторые другие d -альдогексозы являются левовращающими.
Превращение между двумя аномерами можно наблюдать в поляриметре , поскольку чистая α- d -глюкоза имеет удельный угол вращения +112,2° мл/(дм·г), чистая β- d -глюкоза +17,5° мл/(дм·г). [63] Когда равновесие достигается через определенное время из-за мутаротации, угол вращения составляет +52,7° мл/(дм·г). [63] При добавлении кислоты или основания это превращение значительно ускоряется. Уравновешивание происходит через альдегидную форму с открытой цепью.
В разбавленном гидроксиде натрия или других разбавленных основаниях моносахариды манноза , глюкоза и фруктоза взаимопревращаются (через преобразование Лобри де Брюйна–Альберды–Ван Экенштейна ), так что между этими изомерами образуется равновесие. Эта реакция протекает через энедиол :

Глюкоза является наиболее распространенным моносахаридом. Глюкоза также является наиболее широко используемой альдогексозой в большинстве живых организмов. Одним из возможных объяснений этого является то, что глюкоза имеет меньшую тенденцию, чем другие альдогексозы, неспецифически реагировать с аминогруппами белков . [ 64] Эта реакция — гликирование — ухудшает или разрушает функцию многих белков, [ 64] например, в гликированном гемоглобине . Низкая скорость гликирования глюкозы может быть связана с тем, что она имеет более стабильную циклическую форму по сравнению с другими альдогексозами, что означает, что она проводит меньше времени, чем они, в своей реактивной форме с открытой цепью. [64] Причина, по которой глюкоза имеет самую стабильную циклическую форму из всех альдогексоз, заключается в том, что ее гидроксильные группы (за исключением гидроксильной группы на аномерном углероде d -глюкозы) находятся в экваториальном положении . Предположительно, глюкоза является наиболее распространенным природным моносахаридом, поскольку она менее гликирована белками, чем другие моносахариды. [64] [65] Другая гипотеза заключается в том, что глюкоза, будучи единственной d -альдогексозой, которая имеет все пять гидроксизаместителей в экваториальном положении в форме β- d -глюкозы, более доступна для химических реакций, [66] : 194, 199 например, для этерификации [67] : 363 или образования ацеталя . [68] По этой причине d -глюкоза также является весьма предпочтительным строительным блоком в природных полисахаридах (гликанах). Полисахариды, которые состоят исключительно из глюкозы, называются глюканами .
Глюкоза вырабатывается растениями посредством фотосинтеза с использованием солнечного света, [69] [70] воды и углекислого газа и может использоваться всеми живыми организмами в качестве источника энергии и углерода. Однако большая часть глюкозы встречается не в свободной форме, а в форме ее полимеров, то есть лактозы, сахарозы, крахмала и других, которые являются энергетическими резервными веществами, а также целлюлозы и хитина , которые являются компонентами клеточной стенки растений или грибов и членистоногих соответственно. Эти полимеры при потреблении животными, грибами и бактериями расщепляются до глюкозы с помощью ферментов. Все животные также способны сами вырабатывать глюкозу из определенных предшественников по мере необходимости. Нейроны , клетки мозгового вещества почек и эритроциты зависят от глюкозы для выработки энергии. [70] У взрослых людей содержится около 18 г (0,63 унции) глюкозы, [71] из которых около 4 г (0,14 унции) присутствует в крови. [72] Примерно 180–220 г (6,3–7,8 унций) глюкозы вырабатывается в печени взрослого человека за 24 часа. [71]
Многие из долгосрочных осложнений диабета (например, слепота , почечная недостаточность и периферическая невропатия ), вероятно, вызваны гликированием белков или липидов . [73] Напротив, регулируемое ферментами добавление сахаров к белку называется гликозилированием и имеет важное значение для функционирования многих белков. [74]
Поглощенная глюкоза изначально связывается с рецептором сладкого вкуса на языке у людей. Этот комплекс белков T1R2 и T1R3 позволяет идентифицировать источники пищи, содержащие глюкозу. [75] [76] Глюкоза в основном поступает с пищей — около 300 г (11 унций) в день производится путем переработки пищи, [76] но она также синтезируется из других метаболитов в клетках организма. У людей расщепление полисахаридов, содержащих глюкозу, происходит частично уже во время жевания с помощью амилазы , которая содержится в слюне , а также мальтазы , лактазы и сахаразы на щеточной каемке тонкого кишечника . Глюкоза является строительным блоком многих углеводов и может быть отщеплена от них с помощью определенных ферментов. Глюкозидазы , подгруппа гликозидаз, сначала катализируют гидролиз длинноцепочечных полисахаридов, содержащих глюкозу, удаляя терминальную глюкозу. В свою очередь, дисахариды в основном расщепляются специфическими гликозидазами до глюкозы. Названия расщепляющих ферментов часто происходят от конкретного поли- и дисахарида; в частности, для расщепления полисахаридных цепей существуют амилазы (названные в честь амилозы, компонента крахмала), целлюлазы (названные в честь целлюлозы), хитиназы (названные в честь хитина) и другие. Кроме того, для расщепления дисахаридов существуют мальтаза, лактаза, сахараза, трегалаза и другие. У людей известно около 70 генов, кодирующих гликозидазы. Они выполняют функции в переваривании и расщеплении гликогена, сфинголипидов , мукополисахаридов и поли( АДФ-рибозы ). Люди не вырабатывают целлюлазы, хитиназы или трегалазы, но бактерии в микробиоте кишечника вырабатывают.
Для того чтобы попасть в клеточные мембраны клеток и мембраны клеточных компартментов или выйти из них, глюкозе требуются специальные транспортные белки из главного суперсемейства посредников . В тонком кишечнике (точнее, в тощей кишке ) [77] глюкоза поступает в эпителий кишечника с помощью транспортеров глюкозы [78] через вторичный активный транспортный механизм, называемый симпортом ионов натрия и глюкозы через котранспортер натрия/глюкозы 1 (SGLT1). [79] Дальнейший перенос происходит на базолатеральной стороне эпителиальных клеток кишечника с помощью транспортера глюкозы GLUT2 [79] , а также поглощение клетками печени , почечными клетками, клетками островков Лангерганса , нейронами , астроцитами и таницитами . [80] Глюкоза поступает в печень через воротную вену и хранится там в виде клеточного гликогена. [81] В клетке печени он фосфорилируется глюкокиназой в позиции 6 с образованием глюкозо-6-фосфата , который не может покинуть клетку. Глюкозо-6-фосфатаза может преобразовывать глюкозо-6-фосфат обратно в глюкозу исключительно в печени, поэтому организм может поддерживать достаточную концентрацию глюкозы в крови. В других клетках поглощение происходит путем пассивного транспорта через один из 14 белков GLUT. [79] В других типах клеток фосфорилирование происходит через гексокиназу , после чего глюкоза больше не может диффундировать из клетки.
Транспортер глюкозы GLUT1 вырабатывается большинством типов клеток и имеет особое значение для нервных клеток и β-клеток поджелудочной железы . [79] GLUT3 высоко экспрессируется в нервных клетках. [79] Глюкоза из кровотока поглощается GLUT4 из мышечных клеток (скелетных мышц [82] и сердечной мышцы ) и жировых клеток . [83] GLUT14 экспрессируется исключительно в яичках . [84] Избыток глюкозы расщепляется и преобразуется в жирные кислоты, которые хранятся в виде триглицеридов . В почках глюкоза в моче поглощается через SGLT1 и SGLT2 в апикальных клеточных мембранах и передается через GLUT2 в базолатеральных клеточных мембранах. [85] Около 90% реабсорбции глюкозы почками происходит через SGLT2 и около 3% через SGLT1. [86]
В растениях и некоторых прокариотах глюкоза является продуктом фотосинтеза . [69] Глюкоза также образуется при распаде полимерных форм глюкозы, таких как гликоген (у животных и грибов ) или крахмал (у растений). Расщепление гликогена называется гликогенолизом, расщепление крахмала называется деградацией крахмала. [87]
Метаболический путь, который начинается с молекул, содержащих от двух до четырех атомов углерода (C), и заканчивается молекулой глюкозы, содержащей шесть атомов углерода, называется глюконеогенезом и встречается во всех живых организмах. Меньшие исходные материалы являются результатом других метаболических путей. В конечном итоге почти все биомолекулы возникают в результате усвоения углекислого газа растениями и микробами во время фотосинтеза. [67] : 359 Свободная энергия образования α- d -глюкозы составляет 917,2 килоджоулей на моль. [67] : 59 У людей глюконеогенез происходит в печени и почках, [88] но также и в других типах клеток. В печени хранится около 150 г (5,3 унции) гликогена, в скелетных мышцах около 250 г (8,8 унции). [89] Однако глюкоза, высвобождаемая в мышечных клетках при расщеплении гликогена, не может быть доставлена в кровоток, поскольку глюкоза фосфорилируется гексокиназой, а глюкозо-6-фосфатаза не экспрессируется для удаления фосфатной группы. В отличие от глюкозы, для глюкозо-6-фосфата нет транспортного белка . Глюконеогенез позволяет организму накапливать глюкозу из других метаболитов, включая лактат или определенные аминокислоты , потребляя при этом энергию. Клетки почечных канальцев также могут вырабатывать глюкозу.
Глюкозу также можно обнаружить вне живых организмов в окружающей среде. Концентрации глюкозы в атмосфере определяются путем сбора образцов самолетами и, как известно, варьируются от места к месту. Например, концентрации глюкозы в атмосферном воздухе из внутреннего Китая колеблются от 0,8 до 20,1 пг/л, тогда как концентрации глюкозы на восточном побережье Китая колеблются от 10,3 до 142 пг/л. [90]
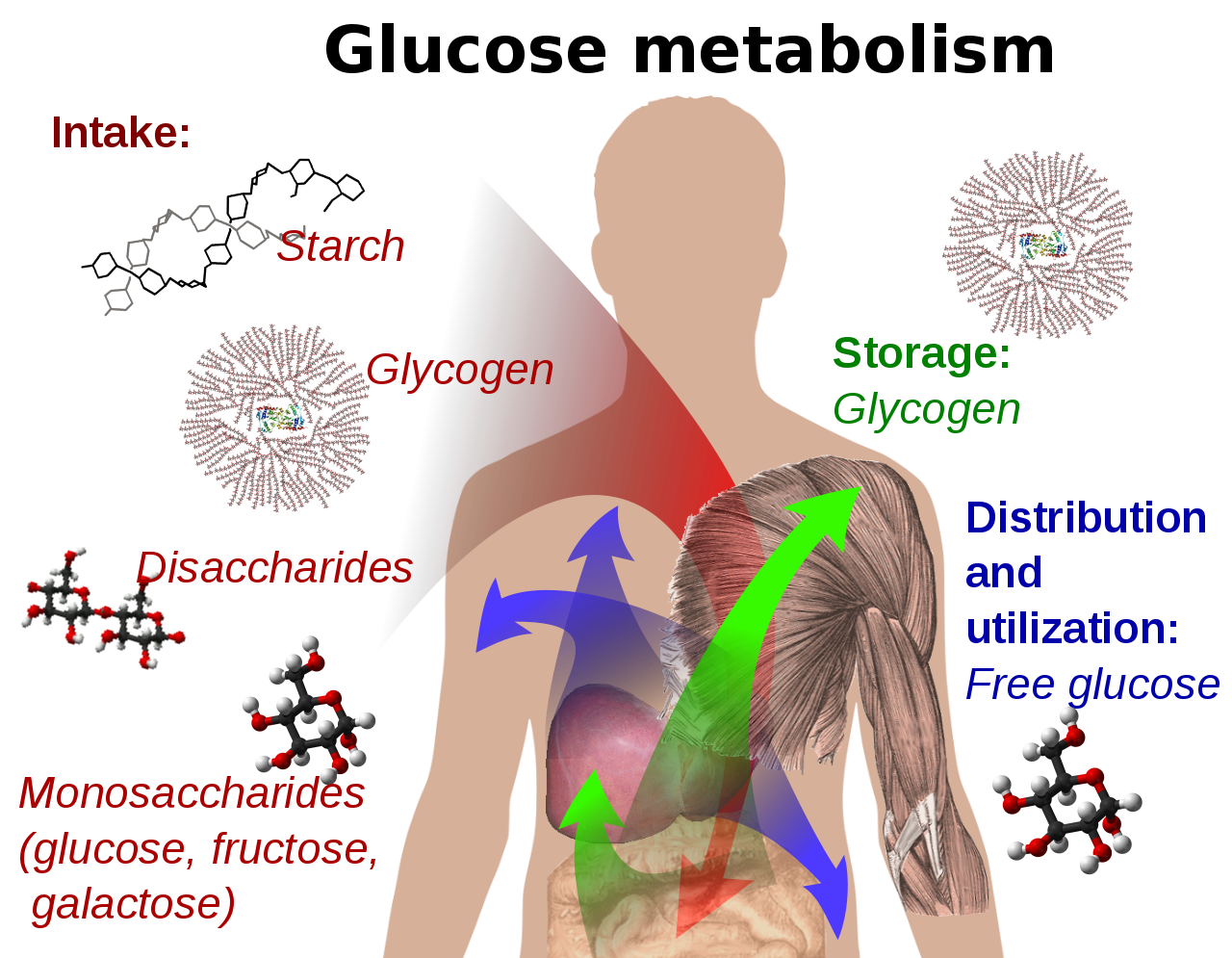
У людей глюкоза метаболизируется посредством гликолиза [91] и пентозофосфатного пути. [92] Гликолиз используется всеми живыми организмами, [66] : 551 [93] с небольшими вариациями, и все организмы вырабатывают энергию из распада моносахаридов. [93] В дальнейшем ходе метаболизма она может быть полностью разрушена посредством окислительного декарбоксилирования , цикла лимонной кислоты (синоним цикла Кребса ) и дыхательной цепи до воды и углекислого газа. Если для этого недостаточно кислорода, расщепление глюкозы у животных происходит анаэробно до лактата через брожение молочной кислоты и выделяет гораздо меньше энергии. Мышечный лактат поступает в печень через кровоток у млекопитающих, где происходит глюконеогенез ( цикл Кори ). При высоком поступлении глюкозы метаболит ацетил-КоА из цикла Кребса также может быть использован для синтеза жирных кислот . [94] Глюкоза также используется для пополнения запасов гликогена в организме, которые в основном находятся в печени и скелетных мышцах. Эти процессы регулируются гормонально .
В других живых организмах могут происходить и другие формы ферментации. Бактерия Escherichia coli может расти на питательных средах, содержащих глюкозу в качестве единственного источника углерода. [67] : 59 У некоторых бактерий и, в измененной форме, также у архей, глюкоза расщепляется по пути Энтнера-Дудорова . [95] С глюкозой был обнаружен механизм регуляции генов в E. coli , катаболитная репрессия (ранее известная как эффект глюкозы ). [96]
Использование глюкозы в качестве источника энергии в клетках происходит либо посредством аэробного дыхания, либо посредством анаэробного дыхания, либо посредством ферментации. [97] Первым этапом гликолиза является фосфорилирование глюкозы гексокиназой с образованием глюкозо-6-фосфата . Основной причиной немедленного фосфорилирования глюкозы является предотвращение ее диффузии из клетки, поскольку заряженная фосфатная группа не позволяет глюкозо-6-фосфату легко пересекать клеточную мембрану . [97] Кроме того, добавление высокоэнергетической фосфатной группы активирует глюкозу для последующего расщепления на более поздних этапах гликолиза. [98]
При анаэробном дыхании одна молекула глюкозы производит чистый прирост в две молекулы АТФ (четыре молекулы АТФ производятся во время гликолиза посредством фосфорилирования на уровне субстрата, но две требуются ферментам, используемым в этом процессе). [99] При аэробном дыхании молекула глюкозы гораздо более выгодна, поскольку генерируется максимальный чистый прирост в 30 или 32 молекулы АТФ (в зависимости от организма). [100]
Нажмите на гены, белки и метаболиты ниже, чтобы перейти к соответствующим статьям. [§ 1]
Опухолевые клетки часто растут сравнительно быстро и потребляют больше среднего количества глюкозы путем гликолиза, [101] что приводит к образованию лактата, конечного продукта ферментации у млекопитающих, даже в присутствии кислорода. Это называется эффектом Варбурга . Для повышенного поглощения глюкозы в опухолях чрезмерно вырабатываются различные SGLT и GLUT. [102] [103]
В дрожжах этанол ферментируется при высоких концентрациях глюкозы, даже в присутствии кислорода (что обычно приводит к дыханию, а не брожению). Это называется эффектом Крэбтри .
Глюкоза также может распадаться с образованием углекислого газа абиотическими способами. Это было продемонстрировано экспериментально путем окисления и гидролиза при 22 °C и pH 2,5. [104]

Глюкоза является вездесущим топливом в биологии . Она используется в качестве источника энергии в организмах, от бактерий до людей, посредством аэробного дыхания , анаэробного дыхания (у бактерий) или ферментации . Глюкоза является основным источником энергии для человеческого организма посредством аэробного дыхания, обеспечивая около 3,75 килокалорий (16 килоджоулей ) пищевой энергии на грамм. [105] Расщепление углеводов (например, крахмала) дает моно- и дисахариды , большую часть которых составляет глюкоза. Через гликолиз и позже в реакциях цикла лимонной кислоты и окислительного фосфорилирования глюкоза окисляется , в конечном итоге образуя диоксид углерода и воду, выделяя энергию в основном в форме аденозинтрифосфата (АТФ). Реакция инсулина и другие механизмы регулируют концентрацию глюкозы в крови. Физиологическая калорийность глюкозы, в зависимости от источника, составляет 16,2 кДж/г [106] или 15,7 кДж/г (3,74 ккал/г). [107] Высокая доступность углеводов из растительной биомассы привела к появлению различных методов в ходе эволюции, особенно у микроорганизмов, для использования глюкозы для получения энергии и хранения углерода. Существуют различия в том, какой конечный продукт больше не может использоваться для производства энергии. Наличие отдельных генов и их генных продуктов, ферментов, определяет, какие реакции возможны. Метаболический путь гликолиза используется почти всеми живыми существами. Существенным отличием в использовании гликолиза является восстановление НАДФН в качестве восстановителя для анаболизма , который в противном случае пришлось бы генерировать косвенно. [108]
Глюкоза и кислород поставляют почти всю энергию для мозга , [109] поэтому ее доступность влияет на психологические процессы. Когда уровень глюкозы низкий , психологические процессы, требующие умственных усилий (например, самоконтроль , принятие трудных решений), нарушаются. [110] [111] [112] [113] В мозге, который зависит от глюкозы и кислорода как основного источника энергии, концентрация глюкозы обычно составляет от 4 до 6 мМ (5 мМ равны 90 мг/дл), [71] но снижается до 2-3 мМ при голодании. [114] Спутанность сознания возникает ниже 1 мМ, а кома - при более низких уровнях. [114]
Глюкоза в крови называется сахаром крови . Уровень сахара в крови регулируется нервными клетками, связывающими глюкозу, в гипоталамусе . [115] Кроме того, глюкоза в мозге связывается с рецепторами глюкозы системы вознаграждения в прилежащем ядре . [115] Связывание глюкозы с рецептором сладкого на языке вызывает высвобождение различных гормонов энергетического обмена, либо через глюкозу, либо через другие сахара, что приводит к повышенному усвоению клетками и снижению уровня сахара в крови. [116] Искусственные подсластители не снижают уровень сахара в крови. [116]
Содержание сахара в крови здорового человека в состоянии кратковременного голодания, например, после ночного голодания, составляет около 70–100 мг/дл крови (4–5,5 мМ). В плазме крови измеренные значения примерно на 10–15% выше. Кроме того, значения в артериальной крови выше, чем концентрации в венозной крови, поскольку глюкоза всасывается в ткани во время прохождения капиллярного русла . Также в капиллярной крови, которая часто используется для определения сахара в крови, значения иногда выше, чем в венозной крови. Содержание глюкозы в крови регулируется гормонами инсулином , инкретином и глюкагоном . [115] [117] Инсулин снижает уровень глюкозы, глюкагон повышает его. [71] Кроме того, гормоны адреналин , тироксин , глюкокортикоиды , соматотропин и адренокортикотропин приводят к повышению уровня глюкозы. [71] Существует также гормононезависимая регуляция, которая называется ауторегуляцией глюкозы. [118] После приема пищи концентрация сахара в крови увеличивается. Значения более 180 мг/дл в венозной цельной крови являются патологическими и называются гипергликемией , значения ниже 40 мг/дл называются гипогликемией . [119] При необходимости глюкоза высвобождается в кровоток глюкозо-6-фосфатазой из глюкозо-6-фосфата, происходящего из гликогена печени и почек, тем самым регулируя гомеостаз концентрации глюкозы в крови. [88] [70] У жвачных животных концентрация глюкозы в крови ниже (60 мг/дл у крупного рогатого скота и 40 мг/дл у овец ), потому что углеводы больше преобразуются их кишечной микробиотой в короткоцепочечные жирные кислоты . [120]
Часть глюкозы преобразуется в молочную кислоту астроцитами , которая затем используется в качестве источника энергии клетками мозга ; часть глюкозы используется кишечными клетками и эритроцитами , в то время как остальная часть достигает печени , жировой ткани и мышечных клеток, где она поглощается и хранится в виде гликогена (под влиянием инсулина ). Гликоген клеток печени может быть преобразован в глюкозу и возвращен в кровь, когда инсулин низкий или отсутствует; гликоген мышечных клеток не возвращается в кровь из-за недостатка ферментов. В жировых клетках глюкоза используется для питания реакций, которые синтезируют некоторые типы жиров и имеют другие цели. Гликоген является механизмом «хранения энергии глюкозы» организма, потому что он гораздо более «экономичен в пространстве» и менее реактивен, чем сама глюкоза.
В связи с ее важностью для здоровья человека глюкоза является аналитом в анализах на глюкозу , которые являются обычными медицинскими анализами крови . [121] Прием пищи или голодание перед взятием образца крови оказывают влияние на анализы на глюкозу в крови; высокий уровень сахара в крови натощак может быть признаком преддиабета или сахарного диабета . [122]
Гликемический индекс является показателем скорости резорбции и преобразования в уровень глюкозы в крови из потребленных углеводов, измеряется как площадь под кривой уровня глюкозы в крови после потребления по сравнению с глюкозой (глюкоза определяется как 100). [123] Клиническое значение гликемического индекса является спорным, [123] [124] поскольку продукты с высоким содержанием жира замедляют резорбцию углеводов и снижают гликемический индекс, например, мороженое. [124] Альтернативным показателем является инсулиновый индекс , [125] измеряемый как влияние потребления углеводов на уровень инсулина в крови. Гликемическая нагрузка является показателем количества глюкозы, добавленной к уровню глюкозы в крови после потребления, на основе гликемического индекса и количества потребленной пищи.
Организмы используют глюкозу в качестве предшественника для синтеза нескольких важных веществ. Крахмал, целлюлоза и гликоген («животный крахмал») являются распространенными полимерами глюкозы (полисахаридами). Некоторые из этих полимеров (крахмал или гликоген) служат хранилищами энергии, в то время как другие (целлюлоза и хитин , который производится из производного глюкозы) играют структурную роль. Олигосахариды глюкозы в сочетании с другими сахарами служат важными хранилищами энергии. К ним относятся лактоза, преобладающий сахар в молоке, который является дисахаридом глюкозы-галактозы, и сахароза, другой дисахарид, который состоит из глюкозы и фруктозы. Глюкоза также добавляется к определенным белкам и липидам в процессе, называемом гликозилированием . Это часто имеет решающее значение для их функционирования. Ферменты, которые присоединяют глюкозу к другим молекулам, обычно используют фосфорилированную глюкозу для питания образования новой связи, соединяя ее с разрывом связи глюкозы-фосфата.
Помимо прямого использования в качестве мономера, глюкоза может расщепляться для синтеза большого количества других биомолекул. Это важно, поскольку глюкоза служит как первичным хранилищем энергии, так и источником органического углерода. Глюкоза может расщепляться и превращаться в липиды. Она также является предшественником синтеза других важных молекул, таких как витамин С (аскорбиновая кислота). В живых организмах глюкоза преобразуется в несколько других химических соединений, которые являются исходным материалом для различных метаболических путей . Среди них все другие моносахариды [126], такие как фруктоза (через полиоловый путь ), [79] манноза (эпимер глюкозы в положении 2), галактоза (эпимер в положении 4), фукоза, различные уроновые кислоты и аминосахара , производятся из глюкозы. [81] В дополнение к фосфорилированию до глюкозо-6-фосфата, которое является частью гликолиза, глюкоза может окисляться во время ее распада до глюконо-1,5-лактона. Глюкоза используется некоторыми бактериями в качестве строительного блока в биосинтезе трегалозы или декстрана , а у животных — в качестве строительного блока гликогена. Глюкоза также может быть преобразована из бактериальной ксилозоизомеразы во фруктозу. Кроме того, метаболиты глюкозы производят все заменимые аминокислоты, сахарные спирты, такие как маннит и сорбит , жирные кислоты , холестерин и нуклеиновые кислоты . [126] Наконец, глюкоза используется в качестве строительного блока в гликозилировании белков в гликопротеины , гликолипиды , пептидогликаны , гликозиды и другие вещества (катализируемом гликозилтрансферазами ) и может быть отщеплена от них гликозидазами .
Диабет — это нарушение обмена веществ, при котором организм не может регулировать уровень глюкозы в крови либо из-за недостатка инсулина в организме, либо из-за неспособности клеток организма должным образом реагировать на инсулин. Каждая из этих ситуаций может быть вызвана постоянно высоким уровнем глюкозы в крови из-за истощения поджелудочной железы и резистентности к инсулину . Поджелудочная железа — это орган, отвечающий за секрецию гормонов инсулина и глюкагона. [127] Инсулин — это гормон, который регулирует уровень глюкозы, позволяя клеткам организма поглощать и использовать глюкозу. Без него глюкоза не может проникать в клетку и, следовательно, не может использоваться в качестве топлива для функций организма. [128] Если поджелудочная железа подвергается постоянно высокому уровню глюкозы в крови, клетки, вырабатывающие инсулин, в поджелудочной железе могут быть повреждены, что приведет к недостатку инсулина в организме. Инсулинорезистентность возникает, когда поджелудочная железа пытается вырабатывать все больше и больше инсулина в ответ на постоянно повышенный уровень глюкозы в крови. В конце концов, остальная часть тела становится устойчивой к инсулину, который вырабатывает поджелудочная железа, тем самым требуя больше инсулина для достижения того же эффекта снижения уровня глюкозы в крови, и заставляя поджелудочную железу вырабатывать еще больше инсулина, чтобы конкурировать с устойчивостью. Эта негативная спираль способствует выгоранию поджелудочной железы и прогрессированию заболевания диабетом.
Чтобы контролировать реакцию организма на терапию, снижающую уровень глюкозы в крови, можно измерить уровень глюкозы. Мониторинг уровня глюкозы в крови можно проводить несколькими методами, например, тестом на глюкозу натощак, который измеряет уровень глюкозы в крови после 8 часов голодания. Другим тестом является 2-часовой тест на толерантность к глюкозе (GTT) — для этого теста человек проходит тест на глюкозу натощак, затем выпивает 75-граммовый напиток с глюкозой и проходит повторное тестирование. Этот тест измеряет способность организма человека перерабатывать глюкозу. Со временем уровень глюкозы в крови должен снижаться, поскольку инсулин позволяет ей поглощаться клетками и выходить из кровотока.

Люди с диабетом или другими заболеваниями, которые приводят к низкому уровню сахара в крови , часто носят с собой небольшое количество сахара в различных формах. Одним из часто используемых видов сахара является глюкоза, часто в форме таблеток глюкозы (глюкоза, спрессованная в форму таблетки, иногда с одним или несколькими другими ингредиентами в качестве связующего вещества), леденцов или сахарных пакетиков .

Большинство пищевых углеводов содержат глюкозу, либо как единственный строительный блок (как в полисахаридах крахмале и гликогене), либо вместе с другим моносахаридом (как в гетерополисахаридах сахарозе и лактозе). [129] Несвязанная глюкоза является одним из основных ингредиентов меда. Глюкоза чрезвычайно распространена и была выделена из различных природных источников по всему миру, включая мужские шишки хвойного дерева Wollemia nobilis в Риме, [130] корни растений Ilex asprella в Китае, [131] и соломку из риса в Калифорнии. [132]
Глюкоза производится промышленным способом из крахмала путем ферментативного гидролиза с использованием глюкозоамилазы или с использованием кислот . Ферментативный гидролиз в значительной степени вытеснил реакции кислотно-катализируемого гидролиза. [134] Результатом является глюкозный сироп (ферментативно с более чем 90% глюкозы в сухом веществе) [134] с годовым мировым объемом производства 20 миллионов тонн (по состоянию на 2011 год). [135] Это причина прежнего распространенного названия «крахмальный сахар». Амилазы чаще всего происходят из Bacillus licheniformis [136] или Bacillus subtilis (штамм MN-385), [136] которые более термостабильны, чем первоначально используемые ферменты. [136] [137] Начиная с 1982 года, пуллуланазы из Aspergillus niger использовались в производстве глюкозного сиропа для преобразования амилопектина в крахмал (амилозу), тем самым увеличивая выход глюкозы. [138] Реакция проводится при pH = 4,6–5,2 и температуре 55–60 °C. [11] Кукурузный сироп содержит от 20% до 95% глюкозы в сухом веществе. [139] [140] Японская форма глюкозного сиропа, Mizuame , производится из батата или рисового крахмала. [141] Мальтодекстрин содержит около 20% глюкозы.
Многие культуры могут быть использованы в качестве источника крахмала. Кукуруза , [134] рис, [134] пшеница , [134] маниока , [134] картофель , [134] ячмень , [134] батат , [142] кукурузная шелуха и саго используются в разных частях мира. В Соединенных Штатах кукурузный крахмал (из кукурузы) используется почти исключительно. Некоторая коммерческая глюкоза встречается как компонент инвертного сахара , примерно 1:1 смеси глюкозы и фруктозы, которая производится из сахарозы. В принципе, целлюлозу можно гидролизовать до глюкозы, но этот процесс пока не является коммерчески практичным. [54]
В США в качестве источника глюкозы для производства изоглюкозы , представляющей собой смесь глюкозы и фруктозы, используется почти исключительно кукуруза (точнее, кукурузный сироп), поскольку фруктоза обладает более высокой подслащивающей способностью — при той же физиологической калорийности 374 килокалории на 100 г. Ежегодное мировое производство изоглюкозы составляет 8 миллионов тонн (по состоянию на 2011 год). [135] При производстве из кукурузного сиропа конечный продукт представляет собой кукурузный сироп с высоким содержанием фруктозы (HFCS).

Глюкоза в основном используется для производства фруктозы и содержащих глюкозу продуктов. В пищевых продуктах она используется как подсластитель, увлажнитель , для увеличения объема и создания более мягкого вкуса . [134] Различные источники глюкозы, такие как виноградный сок (для вина) или солод (для пива), используются для ферментации в этанол во время производства алкогольных напитков . Большинство безалкогольных напитков в США используют HFCS-55 (с содержанием фруктозы 55% в сухой массе), в то время как большинство других подслащенных HFCS продуктов в США используют HFCS-42 (с содержанием фруктозы 42% в сухой массе). [144] В Мексике, с другой стороны, безалкогольные напитки подслащивают тростниковым сахаром, который имеет более высокую подслащивающую способность. [145] Кроме того, глюкозный сироп используется, в частности, в производстве кондитерских изделий , таких как конфеты , ириски и помадки . [146] Типичными химическими реакциями глюкозы при нагревании в условиях отсутствия воды являются карамелизация и, в присутствии аминокислот, реакция Майяра .
Кроме того, из глюкозы биотехнологическим путем можно получать различные органические кислоты, например, путем ферментации с помощью Clostridium thermoaceticum для получения уксусной кислоты , с помощью Penicillium notatum для получения арабоаскорбиновой кислоты, с помощью Rhizopus delemar для получения фумаровой кислоты , с помощью Aspergillus niger для получения глюконовой кислоты , с помощью Candida brumptii для получения изолимонной кислоты , с помощью Aspergillus terreus для получения итаконовой кислоты , с помощью Pseudomonas fluorescens для получения 2-кетоглюконовой кислоты, с помощью Gluconobacter suboxydans для получения 5-кетоглюконовой кислоты, с помощью Aspergillus oryzae для получения койевой кислоты , с помощью Lactobacillus delbrueckii для получения молочной кислоты , с помощью Lactobacillus brevis для получения яблочной кислоты , с помощью Propionibacter shermanii для производства пропионовой кислоты , с Pseudomonas aeruginosa для производства пировиноградной кислоты и с Gluconobacter suboxydans для производства винной кислоты . [147] [ необходимы дополнительные ссылки ] Недавно сообщалось о мощных биоактивных натуральных продуктах, таких как триптолид, которые ингибируют транскрипцию млекопитающих посредством ингибирования субъединицы XPB общего фактора транскрипции TFIIH, как о конъюгате глюкозы для нацеливания на гипоксические раковые клетки с повышенной экспрессией транспортера глюкозы. [148] В последнее время глюкоза получает коммерческое использование в качестве ключевого компонента «наборов», содержащих молочную кислоту и инсулин, предназначенных для индукции гипогликемии и гиперлактатемии для борьбы с различными видами рака и инфекциями. [149]
Когда необходимо обнаружить молекулу глюкозы в определенном положении в более крупной молекуле, проводится ядерно-магнитная резонансная спектроскопия , рентгеноструктурный анализ или иммуноокрашивание лектина с использованием конъюгата фермента-репортера конканавалина А , который связывает только глюкозу или маннозу.
Эти реакции имеют только историческое значение:
Тест Фелинга является классическим методом обнаружения альдоз. [150] Из-за мутаротации глюкоза всегда присутствует в небольшом количестве в виде альдегида с открытой цепью. При добавлении реагентов Фелинга (раствор Фелинга (I) и раствор Фелинга (II)) альдегидная группа окисляется до карбоновой кислоты , в то время как комплекс тартрата Cu2 + восстанавливается до Cu + и образует кирпично-красный осадок (Cu2O ) .
В тесте Толленса после добавления аммиачного AgNO 3 к раствору образца глюкоза восстанавливает Ag + до элементарного серебра . [151]
В тесте Барфоеда [152] раствор растворенного ацетата меди , ацетата натрия и уксусной кислоты добавляется к раствору сахара, который необходимо проверить, и затем нагревается на водяной бане в течение нескольких минут. Глюкоза и другие моносахариды быстро дают красноватый цвет и красновато-коричневый оксид меди(I) (Cu2O ) .
Как восстанавливающий сахар, глюкоза реагирует в тесте Ниландера . [153]
При нагревании разбавленного раствора гидроксида калия с глюкозой до 100 °C появляется сильное красноватое потемнение и карамельный запах. [154] Концентрированная серная кислота растворяет сухую глюкозу без почернения при комнатной температуре, образуя сахарную серную кислоту. [154] [ требуется проверка ] В дрожжевом растворе спиртовое брожение производит углекислый газ в соотношении 2,0454 молекулы глюкозы на одну молекулу CO 2 . [154] Глюкоза образует черную массу с хлоридом олова . [154] В аммиачном растворе серебра глюкоза (а также лактоза и декстрин) приводит к осаждению серебра. В аммиачном растворе ацетата свинца в присутствии глюкозы образуется белый гликозид свинца, который становится менее растворимым при варке и становится коричневым. [154] В аммиачном растворе меди желтый гидрат оксида меди образуется с глюкозой при комнатной температуре, в то время как красный оксид меди образуется во время кипячения (то же самое с декстрином, за исключением аммиачного раствора ацетата меди). [154] С реагентом Хагера глюкоза образует оксид ртути во время кипячения. [154] Щелочной раствор висмута используется для осаждения элементарного, черно-коричневого висмута с глюкозой. [154] Глюкоза, кипяченная в растворе молибдата аммония, окрашивает раствор в синий цвет. Раствор с индигокармином и карбонатом натрия обесцвечивается при кипячении с глюкозой. [154]
В концентрированных растворах глюкозы с низкой долей других углеводов ее концентрацию можно определить с помощью поляриметра. Для сахарных смесей концентрацию можно определить с помощью рефрактометра , например, при определении Эксле в процессе производства вина.
Фермент глюкозооксидаза (GOx) превращает глюкозу в глюконовую кислоту и перекись водорода, потребляя кислород. Другой фермент, пероксидаза, катализирует хромогенную реакцию (реакция Триндера) [155] фенола с 4-аминоантипирином в пурпурный краситель. [156]
Метод тест-полосок использует вышеупомянутое ферментативное превращение глюкозы в глюконовую кислоту с образованием перекиси водорода. Реагенты иммобилизованы на полимерной матрице, так называемой тест-полоске, которая принимает более или менее интенсивный цвет. Это можно измерить рефлектометрически при 510 нм с помощью ручного фотометра на основе светодиодов. Это позволяет проводить рутинное определение сахара в крови неспециалистам. В дополнение к реакции фенола с 4-аминоантипирином были разработаны новые хромогенные реакции, которые позволяют проводить фотометрию при более высоких длинах волн (550 нм, 750 нм). [156] [157]
Электроанализ глюкозы также основан на ферментативной реакции, упомянутой выше. Полученный пероксид водорода может быть количественно амперометрически определен путем анодного окисления при потенциале 600 мВ. [158] GOx иммобилизован на поверхности электрода или в мембране, расположенной близко к электроду. Драгоценные металлы, такие как платина или золото, используются в электродах, а также электроды из углеродных нанотрубок, которые, например, легированы бором. [159] Нанопроволоки Cu–CuO также используются в качестве амперометрических электродов без ферментов, достигая предела обнаружения 50 мкмоль/л. [160] Особенно многообещающим методом является так называемая «ферментная проводка», где электрон, текущий во время окисления, передается через молекулярный провод непосредственно от фермента к электроду. [161]
Существует множество других химических датчиков для измерения глюкозы. [162] [163] Учитывая важность анализа глюкозы в науках о жизни, также были разработаны многочисленные оптические зонды для сахаридов на основе использования бороновых кислот, [164] которые особенно полезны для внутриклеточных сенсорных приложений, где другие (оптические) методы не применимы или применимы только условно. В дополнение к производным органических бороновых кислот, которые часто связываются с высокой специфичностью с 1,2-диольными группами сахаров, существуют также другие концепции зондов, классифицированные по функциональным механизмам, которые используют селективные связывающие глюкозу белки (например, конканавалин А) в качестве рецептора. Кроме того, были разработаны методы, которые косвенно определяют концентрацию глюкозы через концентрацию метаболизированных продуктов, например, по потреблению кислорода с использованием флуоресцентно-оптических датчиков. [165] Наконец, существуют концепции на основе ферментов, которые используют собственное поглощение или флуоресценцию (флуоресцентно-меченых) ферментов в качестве репортеров. [162]
Глюкозу можно количественно определить с помощью медной йодометрии. [166]
В частности, для анализа сложных смесей, содержащих глюкозу, например, в меде, хроматографические методы, такие как высокоэффективная жидкостная хроматография и газовая хроматография [166], часто используются в сочетании с масс-спектрометрией . [167] [168] Принимая во внимание изотопные соотношения, с помощью этих методов также можно надежно обнаружить фальсификацию меда добавленными сахарами. [169] Обычно используется дериватизация с использованием силилирующих реагентов. [170] Также можно количественно определить пропорции ди- и трисахаридов.
Поглощение глюкозы в клетках организмов измеряется с помощью 2-дезокси-D-глюкозы или фтордезоксиглюкозы . [114] ( 18 F)фтордезоксиглюкоза используется в качестве трассера в позитронно-эмиссионной томографии в онкологии и неврологии, [171] где она является наиболее часто используемым диагностическим агентом. [172]