Биохимия или биологическая химия — это изучение химических процессов внутри живых организмов и связанных с ними . [1] Биохимия , являющаяся субдисциплиной химии и биологии , может быть разделена на три области: структурная биология , энзимология и метаболизм . За последние десятилетия 20-го века биохимия добилась успеха в объяснении жизненных процессов с помощью этих трех дисциплин. Почти все области наук о жизни открываются и развиваются посредством биохимической методологии и исследований. [2] Биохимия фокусируется на понимании химической основы, которая позволяет биологическим молекулам вызывать процессы, происходящие внутри живых клеток и между клетками, [3] , что, в свою очередь, во многом связано с пониманием тканей и органов , а также структуры и функций организма. . [4] Биохимия тесно связана с молекулярной биологией , изучением молекулярных механизмов биологических явлений. [5]
Большая часть биохимии занимается структурами, связями, функциями и взаимодействиями биологических макромолекул , таких как белки , нуклеиновые кислоты , углеводы и липиды . Они обеспечивают структуру клеток и выполняют многие функции, связанные с жизнью. [6] Химия клетки также зависит от реакций малых молекул и ионов . Они могут быть неорганическими (например, вода и ионы металлов ) или органическими (например, аминокислоты , которые используются для синтеза белков ). [7] Механизмы, используемые клетками для использования энергии из окружающей среды посредством химических реакций , известны как метаболизм . Результаты биохимии применяются главным образом в медицине , питании и сельском хозяйстве . В медицине биохимики исследуют причины и способы лечения болезней . [8] Питание изучает способы поддержания здоровья и хорошего самочувствия, а также последствия дефицита питательных веществ . [9] В сельском хозяйстве биохимики исследуют почву и удобрения с целью улучшения выращивания сельскохозяйственных культур, хранения урожая и борьбы с вредителями . В последние десятилетия биохимические принципы и методы были объединены с инженерными подходами к решению проблем , позволяющими манипулировать живыми системами, чтобы создать полезные инструменты для исследований, промышленных процессов, а также диагностики и контроля заболеваний — дисциплины биотехнологии . [10]
_and_Carl_Ferdinand_Cori_-_restoration1.jpg/1280px-Gerty_Theresa_Radnitz_Cori_(1896-1957)_and_Carl_Ferdinand_Cori_-_restoration1.jpg)
В наиболее полном определении биохимию можно рассматривать как исследование компонентов и состава живых существ и того, как они объединяются, образуя жизнь. В этом смысле история биохимии может восходить к древним грекам . [11] Однако биохимия как отдельная научная дисциплина возникла где-то в XIX веке или немного раньше, в зависимости от того, на каком аспекте биохимии основное внимание уделяется. Некоторые утверждали, что началом биохимии могло быть открытие первого фермента , диастазы (ныне называемой амилазой ), в 1833 году Ансельмом Пайеном , [12] в то время как другие считали первую демонстрацию Эдуардом Бюхнером сложного биохимического процесса алкогольного брожения в бесклеточные экстракты в 1897 году стали рождением биохимии. [13] [14] Некоторые могут также указать в качестве начала на влиятельную работу Юстуса фон Либиха 1842 года «Химия животных , или Органическая химия в ее приложениях к физиологии и патологии» , в которой представлена химическая теория метаболизма, [11] или еще раньше, в 18 веке, исследования брожения и дыхания Антуана Лавуазье . [15] [16] Многие другие пионеры в этой области, которые помогли раскрыть уровни сложности биохимии, были провозглашены основателями современной биохимии. Эмиль Фишер , изучавший химию белков , [17] и Ф. Гоуленд Хопкинс , изучавший ферменты и динамическую природу биохимии, представляют собой два примера ранних биохимиков. [18]
Термин «биохимия» впервые был использован, когда Винценц Клецинский (1826–1882) напечатал в Вене в 1858 году свой «Сборник биохимии»; оно получено в результате сочетания биологии и химии . В 1877 году Феликс Хоппе-Зейлер использовал этот термин ( биохимия на немецком языке) как синоним физиологической химии в предисловии к первому выпуску Zeitschrift für Physiologische Chemie (Журнал физиологической химии), где он выступал за создание институтов, посвященных физиологической химии. эту область обучения. [19] [20] Однако часто упоминается, что немецкий химик Карл Нойберг придумал это слово в 1903 году, [21] [22] [23] , в то время как некоторые приписывают его Францу Хофмейстеру . [24]

Когда-то считалось, что жизнь и ее материалы обладают неким существенным свойством или веществом (часто называемым « жизненным принципом »), отличным от любого, обнаруженного в неживой материи, и считалось, что только живые существа могут производить молекулы жизнь. [26] В 1828 году Фридрих Вёлер опубликовал статью о своем удачном синтезе мочевины из цианата калия и сульфата аммония; некоторые считали это прямым ниспровержением витализма и утверждением органической химии . [27] [28] Однако синтез Велера вызвал споры, поскольку некоторые отвергают смерть витализма от его рук. [29] С тех пор биохимия продвинулась вперед, особенно с середины 20-го века, с развитием новых методов, таких как хроматография , рентгеновская дифракция , интерферометрия двойной поляризации , ЯМР-спектроскопия , радиоизотопная маркировка , электронная микроскопия и моделирование молекулярной динамики . Эти методы позволили открыть и детально проанализировать многие молекулы и метаболические пути клетки , такие как гликолиз и цикл Кребса (цикл лимонной кислоты), и привели к пониманию биохимии на молекулярном уровне.
Еще одним важным историческим событием в биохимии является открытие гена и его роли в передаче информации в клетке. В 1950-х годах Джеймс Д. Уотсон , Фрэнсис Крик , Розалинда Франклин и Морис Уилкинс сыграли важную роль в разгадке структуры ДНК и предположили ее связь с генетической передачей информации. [30] В 1958 году Джордж Бидл и Эдвард Татум получили Нобелевскую премию за работу над грибами, показавшую, что один ген производит один фермент . [31] В 1988 году Колин Питчфорк был первым человеком, осужденным за убийство с помощью доказательств ДНК , что привело к развитию судебной медицины . [32] Совсем недавно Эндрю З. Файер и Крейг К. Мелло получили Нобелевскую премию 2006 года за открытие роли РНК-интерференции (РНКи) в подавлении экспрессии генов . [33]

Около двух десятков химических элементов необходимы для различных видов биологической жизни . Большинство редких элементов на Земле не нужны для жизни (исключение составляют селен и йод ), [34] а некоторые распространенные ( алюминий и титан ) не используются. У большинства организмов общие потребности в элементах, но между растениями и животными есть некоторые различия . Например, океанские водоросли используют бром , но наземные растения и животные, похоже, в нем не нуждаются. Всем животным требуется натрий , но для растений он не является важным элементом. Растениям нужны бор и кремний , но животным они могут не потребоваться (или им могут потребоваться очень небольшие количества).
Всего шесть элементов — углерод , водород , азот , кислород , кальций и фосфор — составляют почти 99% массы живых клеток, в том числе и в организме человека ( полный список см. в составе человеческого тела ). В дополнение к шести основным элементам, которые составляют большую часть человеческого тела, людям требуется меньшее количество, возможно, еще 18. [35]
Четыре основных класса молекул в биохимии (часто называемые биомолекулами ) — это углеводы , липиды , белки и нуклеиновые кислоты . [36] Многие биологические молекулы являются полимерами : в этой терминологии мономеры представляют собой относительно небольшие макромолекулы, которые соединяются вместе, образуя большие макромолекулы , известные как полимеры. Когда мономеры соединяются вместе для синтеза биологического полимера , они подвергаются процессу, называемому синтезом дегидратации . Различные макромолекулы могут собираться в более крупные комплексы, часто необходимые для биологической активности .
Двумя основными функциями углеводов являются накопление энергии и обеспечение структуры. Один из распространенных сахаров , известный как глюкоза , является углеводом, но не все углеводы являются сахарами. На Земле больше углеводов, чем любого другого известного типа биомолекул; они используются для хранения энергии и генетической информации , а также играют важную роль в межклеточных взаимодействиях и коммуникациях . [ нужна цитата ]
Простейшим видом углеводов является моносахарид , который помимо других свойств содержит углерод , водород и кислород , преимущественно в соотношении 1:2:1 (обобщенная формула C n H 2 n O n , где n равно не менее 3). Глюкоза (C 6 H 12 O 6 ) — один из важнейших углеводов; другие включают фруктозу (C 6 H 12 O 6 ), сахар, обычно связанный со сладким вкусом фруктов , [37] [ a] и дезоксирибозу (C 5 H 10 O 4 ), компонент ДНК . Моносахарид может переключаться между ациклической (с открытой цепью) формой и циклической формой. Форма с открытой цепью может быть превращена в кольцо атомов углерода, соединенных мостиком атома кислорода , созданного из карбонильной группы на одном конце и гидроксильной группы на другом. Циклическая молекула имеет гемиацетальную или гемикетальную группу, в зависимости от того, была ли линейная форма альдозой или кетозой . [38]
В этих циклических формах кольцо обычно имеет 5 или 6 атомов. Эти формы называются фуранозами и пиранозами соответственно — по аналогии с фураном и пираном , простейшими соединениями с одинаковым углерод-кислородным кольцом (хотя у них отсутствуют двойные углерод-углеродные связи этих двух молекул). Например, альдогексоза- глюкоза может образовывать полуацетальную связь между гидроксилом на углероде 1 и кислородом на углероде 4, образуя молекулу с 5-членным кольцом, называемую глюкофуранозой . Та же самая реакция может происходить между атомами углерода 1 и 5 с образованием молекулы с 6-членным кольцом, называемой глюкопиранозой . Циклические формы с 7-атомным кольцом, называемые гептозами , встречаются редко. [ нужна цитата ]
Два моносахарида могут быть соединены гликозидной или сложноэфирной связью в дисахарид посредством реакции дегидратации, в ходе которой высвобождается молекула воды. Обратная реакция, при которой гликозидная связь дисахарида разрывается на два моносахарида, называется гидролизом . Самый известный дисахарид — сахароза или обычный сахар , который состоит из соединенных молекул глюкозы и молекулы фруктозы . Другим важным дисахаридом является лактоза , содержащаяся в молоке, состоящая из молекулы глюкозы и молекулы галактозы . Лактоза может гидролизоваться лактазой , а дефицит этого фермента приводит к непереносимости лактозы .
Когда несколько (около трех-шести) моносахаридов соединены, это называется олигосахаридом ( « олиго » означает «немного»). Эти молекулы, как правило, используются в качестве маркеров и сигналов , а также имеют и другие применения. [39] Многие моносахариды, соединенные, образуют полисахарид . Они могут быть соединены в одну длинную линейную цепь, а могут быть и разветвленными . Двумя наиболее распространенными полисахаридами являются целлюлоза и гликоген , оба состоят из повторяющихся мономеров глюкозы . Целлюлоза является важным структурным компонентом клеточных стенок растений, а гликоген используется в качестве хранилища энергии у животных.
Сахар может характеризоваться наличием редуцирующих или невосстанавливающих концов. Восстанавливающий конец углеводов — это атом углерода, который может находиться в равновесии с альдегидом с открытой цепью ( альдоза ) или кето-формой ( кетоза ). Если присоединение мономеров происходит по такому атому углерода, свободная гидроксигруппа пиранозной или фуранозной формы заменяется на ОН-боковую цепь другого сахара, образуя полный ацеталь . Это предотвращает раскрытие цепи в альдегидную или кето-форму и делает модифицированный остаток невосстанавливающим. Лактоза содержит восстанавливающий конец глюкозной части, тогда как галактозная часть образует полный ацеталь с группой C4-OH глюкозы. Сахароза не имеет восстанавливающего конца из-за полного образования ацеталя между альдегидным углеродом глюкозы (С1) и кетоуглеродом фруктозы (С2).

Липиды включают в себя разнообразный набор молекул и в некоторой степени являются совокупностью относительно водонерастворимых или неполярных соединений биологического происхождения, включая воски , жирные кислоты , фосфолипиды , полученные из жирных кислот, сфинголипиды , гликолипиды и терпеноиды (например, ретиноиды и стероиды) . ). Некоторые липиды представляют собой линейные алифатические молекулы с открытой цепью, тогда как другие имеют кольцевую структуру. Некоторые из них являются ароматическими (с циклической [кольцевой] и плоской [плоской] структурой), а другие — нет. Некоторые из них гибкие, а другие жесткие. [42]
Липиды обычно состоят из одной молекулы глицерина в сочетании с другими молекулами. В триглицеридах , основной группе основных липидов, имеется одна молекула глицерина и три жирных кислоты . В этом случае жирные кислоты считаются мономерами и могут быть насыщенными (отсутствие двойных связей в углеродной цепи) или ненасыщенными (одна или несколько двойных связей в углеродной цепи). [ нужна цитата ]
Большинство липидов помимо того, что они в значительной степени неполярны, обладают некоторым полярным характером. Как правило, основная часть их структуры неполярна или гидрофобна («боится воды»), а это означает, что она плохо взаимодействует с полярными растворителями, такими как вода . Другая часть их структуры является полярной или гидрофильной («водолюбивой») и имеет тенденцию связываться с полярными растворителями, такими как вода. Это делает их амфифильными молекулами (имеющими как гидрофобные, так и гидрофильные части). В случае холестерина полярная группа представляет собой просто –ОН (гидроксил или спирт). В случае фосфолипидов полярные группы значительно больше и более полярны, как описано ниже.
Липиды являются неотъемлемой частью нашего ежедневного рациона. Большинство масел и молочных продуктов , которые мы используем для приготовления пищи и еды, таких как сливочное масло , сыр , топленое масло и т. д., состоят из жиров . Растительные масла богаты различными полиненасыщенными жирными кислотами (ПНЖК). Липидосодержащие продукты перевариваются в организме и расщепляются на жирные кислоты и глицерин, которые являются конечными продуктами распада жиров и липидов. Липиды, особенно фосфолипиды , также используются в различных фармацевтических продуктах либо в качестве солюбилизаторов (например, при парентеральных инфузиях), либо в качестве компонентов -носителей лекарственного средства (например, в липосомах или трансферсомах ).
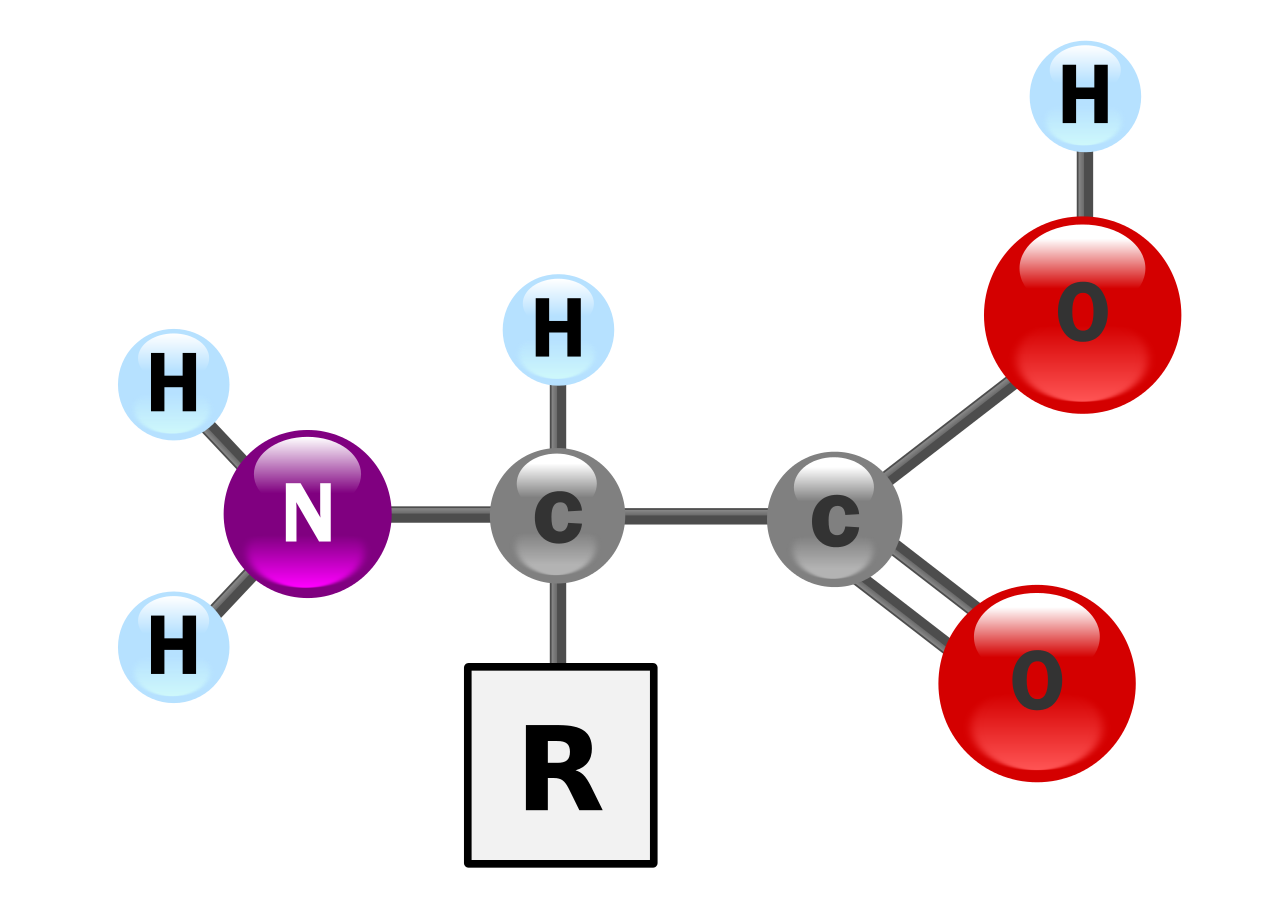
Белки — это очень большие молекулы — макробиополимеры, состоящие из мономеров, называемых аминокислотами . Аминокислота состоит из альфа-атома углерода, присоединенного к аминогруппе –NH 2 , группы карбоновой кислоты –COOH (хотя в физиологических условиях они существуют как –NH 3 + и –COO – ), простого атома водорода и боковая цепь, обычно обозначаемая как «–R». Боковая цепь «R» различна для каждой аминокислоты, из которых имеется 20 стандартных . Именно эта группа «R» делает каждую аминокислоту уникальной, а свойства боковых цепей сильно влияют на общую трехмерную конформацию белка. Некоторые аминокислоты выполняют функции сами по себе или в модифицированной форме; например, глутамат действует как важный нейромедиатор . Аминокислоты могут соединяться посредством пептидной связи . При этом дегидратационном синтезе молекула воды удаляется, и пептидная связь соединяет азот аминогруппы одной аминокислоты с углеродом группы карбоновой кислоты другой. Полученная молекула называется дипептидом , а короткие участки аминокислот (обычно менее тридцати) называются пептидами или полипептидами . Более длинные растяжки заслуживают названия «белки» . Например, важный белок сыворотки крови альбумин содержит 585 аминокислотных остатков . [43]


Белки могут выполнять структурную и/или функциональную роль. Например, движения белков актина и миозина в конечном итоге ответственны за сокращение скелетных мышц. Одним из свойств многих белков является то, что они специфически связываются с определенной молекулой или классом молекул — они могут быть чрезвычайно избирательными в отношении того, с чем связываются. Антитела являются примером белков, которые прикрепляются к одному конкретному типу молекул. Антитела состоят из тяжелых и легких цепей. Две тяжелые цепи будут связаны с двумя легкими цепями посредством дисульфидных связей между их аминокислотами. Антитела специфичны благодаря вариациям, основанным на различиях в N-концевом домене. [44]
Иммуноферментный анализ (ИФА), в котором используются антитела, является одним из наиболее чувствительных тестов, используемых современной медициной для обнаружения различных биомолекул. Однако, вероятно, наиболее важными белками являются ферменты . Практически каждая реакция в живой клетке требует фермента для снижения энергии активации реакции. Эти молекулы распознают специфические молекулы-реагенты, называемые субстратами ; затем они катализируют реакцию между собой. Снижая энергию активации , фермент ускоряет эту реакцию в 10 11 и более раз; реакция, которая обычно спонтанно завершается более 3000 лет, с ферментом может занять менее секунды. Сам фермент не расходуется в процессе и может катализировать ту же реакцию с новым набором субстратов. С помощью различных модификаторов можно регулировать активность фермента, позволяя контролировать биохимию клетки в целом.
Структура белков традиционно описывается в иерархии четырех уровней. Первичная структура белка состоит из линейной последовательности аминокислот; например, «аланин-глицин-триптофан-серин-глутамат-аспарагин-глицин-лизин-...». Вторичная структура связана с локальной морфологией (морфология - это изучение структуры). Некоторые комбинации аминокислот имеют тенденцию сворачиваться в клубок, называемый α-спиралью , или в лист, называемый β-листом ; некоторые α-спирали можно увидеть на схеме гемоглобина выше. Третичная структура – это вся трехмерная форма белка. Эта форма определяется последовательностью аминокислот. Фактически, одно изменение может изменить всю структуру. Альфа-цепь гемоглобина содержит 146 аминокислотных остатков; замена остатка глутамата в положении 6 остатком валина настолько меняет поведение гемоглобина, что приводит к серповидно-клеточной анемии . Наконец, четвертичная структура связана со структурой белка с несколькими пептидными субъединицами, например, гемоглобина с четырьмя субъединицами. Не все белки имеют более одной субъединицы. [45]


Поступившие в организм белки обычно расщепляются на отдельные аминокислоты или дипептиды в тонком кишечнике , а затем всасываются. Затем их можно объединить с образованием новых белков. Промежуточные продукты гликолиза, цикла лимонной кислоты и пентозофосфатного пути могут использоваться для образования всех двадцати аминокислот, а большинство бактерий и растений обладают всеми необходимыми ферментами для их синтеза. Однако люди и другие млекопитающие могут синтезировать только половину из них. Они не могут синтезировать изолейцин , лейцин , лизин , метионин , фенилаланин , треонин , триптофан и валин . Поскольку их необходимо принимать внутрь, это незаменимые аминокислоты . Млекопитающие обладают ферментами для синтеза аланина , аспарагина , аспартата , цистеина , глутамата , глютамина , глицина , пролина , серина и тирозина — заменимых аминокислот. Хотя они могут синтезировать аргинин и гистидин , они не могут производить их в достаточных количествах для молодых растущих животных, поэтому их часто считают незаменимыми аминокислотами.
Если аминогруппа удаляется из аминокислоты, остается углеродный скелет, называемый α- кетокислотой . Ферменты, называемые трансаминазами, могут легко переносить аминогруппу от одной аминокислоты (превращая ее в α-кетокислоту) к другой α-кетокислоте (превращая ее в аминокислоту). Это важно для биосинтеза аминокислот, так как для многих путей промежуточные продукты других биохимических путей преобразуются в скелет α-кетокислоты, а затем добавляется аминогруппа, часто посредством трансаминирования . Аминокислоты затем могут быть связаны вместе с образованием белка.
Аналогичный процесс используется для расщепления белков. Сначала он гидролизуется на составляющие его аминокислоты. Свободный аммиак (NH3), существующий в крови в виде иона аммония (NH4+), токсичен для форм жизни. Поэтому должен существовать подходящий метод его выведения. У разных животных развились разные тактики, в зависимости от их потребностей. Одноклеточные организмы выделяют аммиак в окружающую среду. Точно так же костистые рыбы могут выделять аммиак в воду, где он быстро растворяется. Как правило, млекопитающие преобразуют аммиак в мочевину посредством цикла мочевины .
Чтобы определить, связаны ли два белка или, другими словами, решить, гомологичны они или нет, ученые используют методы сравнения последовательностей. Такие методы, как выравнивание последовательностей и структурное выравнивание, являются мощными инструментами, которые помогают ученым определять гомологию между родственными молекулами. Актуальность поиска гомологии среди белков выходит за рамки формирования эволюционной модели белковых семейств . Выяснив, насколько похожи две белковые последовательности, мы получаем знания об их структуре и, следовательно, об их функциях.

Нуклеиновые кислоты , называемые так из-за их распространенности в клеточных ядрах , являются общим названием семейства биополимеров . Это сложные биохимические макромолекулы с высокой молекулярной массой, которые могут передавать генетическую информацию во всех живых клетках и вирусах. [2] Мономеры называются нуклеотидами , и каждый состоит из трех компонентов: азотистого гетероциклического основания ( пурина или пиримидина ), пентозного сахара и фосфатной группы. [46]

Наиболее распространенными нуклеиновыми кислотами являются дезоксирибонуклеиновая кислота (ДНК) и рибонуклеиновая кислота (РНК). Фосфатная группа и сахар каждого нуклеотида связываются друг с другом, образуя основу нуклеиновой кислоты, а последовательность азотистых оснований хранит информацию. Наиболее распространенными азотистыми основаниями являются аденин , цитозин , гуанин , тимин и урацил . Азотистые основания каждой цепи нуклеиновой кислоты образуют водородные связи с некоторыми другими азотистыми основаниями в комплементарной цепи нуклеиновой кислоты (подобно застежке-молнии). Аденин связывается с тимином и урацилом, тимин связывается только с аденином, а цитозин и гуанин могут связываться только друг с другом. Аденин, тимин, аденин и урацил содержат две водородные связи, тогда как водородных связей, образующихся между цитозином и гуанином, три.
Помимо генетического материала клетки, нуклеиновые кислоты часто играют роль вторичных мессенджеров , а также образуют базовую молекулу аденозинтрифосфата (АТФ), основной молекулы-переносчика энергии, обнаруженной во всех живых организмах. Кроме того, азотистые основания, возможные в двух нуклеиновых кислотах, различны: аденин, цитозин и гуанин встречаются как в РНК, так и в ДНК, тогда как тимин встречается только в ДНК, а урацил встречается в РНК.
Глюкоза является источником энергии у большинства форм жизни. Например, полисахариды расщепляются на мономеры ферментами ( гликогенфосфорилаза удаляет остатки глюкозы из гликогена, полисахарида). Дисахариды, такие как лактоза или сахароза, расщепляются на двухкомпонентные моносахариды. [47]
Глюкоза в основном метаболизируется по очень важному десятиэтапному пути , называемому гликолизом , конечным результатом которого является расщепление одной молекулы глюкозы на две молекулы пирувата . При этом также образуются две чистые молекулы АТФ , энергетической валюты клеток, а также два восстанавливающих эквивалента преобразования НАД + (никотинамидадениндинуклеотид: окисленная форма) в НАДН (никотинамидадениндинуклеотид: восстановленная форма). Для этого не требуется кислород; если кислород недоступен (или клетка не может использовать кислород), НАД восстанавливается путем преобразования пирувата в лактат (молочную кислоту) (например, у людей) или в этанол плюс углекислый газ (например, у дрожжей ). Другие моносахариды, такие как галактоза и фруктоза, могут превращаться в промежуточные продукты гликолитического пути. [48]
В аэробных клетках с достаточным количеством кислорода , как и в большинстве клеток человека, пируват подвергается дальнейшему метаболизму. Он необратимо превращается в ацетил-КоА , выделяя один атом углерода в виде углекислого газа , образуя другой восстанавливающий эквивалент в виде НАДН . Две молекулы ацетил-КоА (из одной молекулы глюкозы) затем вступают в цикл лимонной кислоты , производя две молекулы АТФ, еще шесть молекул НАДН и два восстановленных (уби)хинона (через ФАДН 2 в качестве кофактора, связанного с ферментом) и высвобождая остальные атомы углерода в виде углекислого газа. Образующиеся молекулы НАДН и хинола затем поступают в ферментные комплексы дыхательной цепи, системы переноса электронов , в конечном итоге переносящей электроны на кислород и сохраняющей высвободившуюся энергию в виде градиента протонов над мембраной ( внутренней митохондриальной мембраной у эукариот). Таким образом, кислород восстанавливается до воды и регенерируются исходные акцепторы электронов НАД + и хинон . Вот почему люди вдыхают кислород и выдыхают углекислый газ. Энергия, выделяющаяся при переносе электронов из высокоэнергетических состояний в НАДН и хиноле, сначала сохраняется в виде протонного градиента и преобразуется в АТФ посредством АТФ-синтазы. При этом образуются дополнительные 28 молекул АТФ (24 из 8 НАДН + 4 из 2 хинолов), что в сумме составляет 32 молекулы АТФ, сохраняющихся на распавшуюся глюкозу (две из гликолиза + две из цитратного цикла). [49] Очевидно, что использование кислорода для полного окисления глюкозы дает организму гораздо больше энергии, чем любая независимая от кислорода метаболическая функция, и это считается причиной того, что сложная жизнь появилась только после того, как атмосфера Земли накопила большое количество кислорода.
У позвоночных животных , активно сокращающиеся скелетные мышцы (например, во время поднятия тяжестей или спринтерского бега), не получают достаточного количества кислорода для удовлетворения потребности в энергии, и поэтому они переходят к анаэробному метаболизму , превращая глюкозу в лактат. Комбинация глюкозы неуглеводного происхождения, например, жиров и белков. Это происходит только тогда, когда запасы гликогена в печени истощены. Этот путь представляет собой решающий разворот гликолиза с пирувата на глюкозу и может использовать многие источники, такие как аминокислоты, глицерин и цикл Кребса . Крупномасштабный катаболизм белков и жиров обычно возникает, когда человек страдает от голодания или определенных эндокринных нарушений. [50] Печень регенерирует глюкозу, используя процесс, называемый глюконеогенезом . Этот процесс не совсем противоположен гликолизу и на самом деле требует в три раза больше энергии, получаемой при гликолизе (используется шесть молекул АТФ по сравнению с двумя, полученными при гликолизе). Аналогично вышеуказанным реакциям, полученная глюкоза может затем подвергаться гликолизу в тканях, которым нужна энергия, сохраняться в виде гликогена (или крахмала в растениях) или превращаться в другие моносахариды или присоединяться к ди- или олигосахаридам. Комбинированный путь гликолиза во время физических упражнений, проникновения лактата через кровоток в печень, последующего глюконеогенеза и выброса глюкозы в кровоток называется циклом Кори . [51]

Исследователи в области биохимии используют специфические методы, присущие биохимии, но все чаще сочетают их с методами и идеями, разработанными в области генетики , молекулярной биологии и биофизики . Между этими дисциплинами нет четкой границы. Биохимия изучает химию , необходимую для биологической активности молекул, молекулярная биология изучает их биологическую активность, генетика изучает их наследственность, которая передается их геномом . Это показано на следующей схеме, которая изображает один из возможных вариантов взаимосвязей между полями:
а. ^ Фруктоза — не единственный сахар, содержащийся во фруктах. Глюкоза и сахароза также содержатся в различных количествах в различных фруктах и иногда превышают присутствующую фруктозу. Например, 32% съедобной части фиников составляет глюкоза, по сравнению с 24% фруктозы и 8% сахарозы. Однако персики содержат больше сахарозы (6,66%), чем фруктозы (0,93%) или глюкозы (1,47%). [52]
{{cite book}}: CS1 maint: multiple names: authors list (link){kind=link}